Synteza kwasów tłuszczowych zachodzi w cytoplazmie i retikulum endoplazmatycznym komórki i jest chemicznie podobna do procesu beta-oksydacji, ale z kilkoma kluczowymi różnicami. Pierwsza z nich dotyczy przygotowania substratów do reakcji, w wyniku których powstaje kwas tłuszczowy. Transport acetylo-CoA z mitochondriów ma miejsce, gdy zaczyna się on gromadzić. Dwie cząsteczki mogą odgrywać rolę w przenoszeniu go do cytoplazmy – cytrynian i acetylokarnityna. W wyniku połączenia oksalooctanu z acetylo-CoA w mitochondrium powstaje cytrynian, który przemieszcza się przez błonę, a następnie pod wpływem działania liazy cytrynianowej w cytoplazmie komórki uwalnia acetylo-CoA i oksalooctan. Dodatkowo, gdy wolny acetylo-CoA gromadzi się w mitochondrium, może połączyć się z karnityną i zostać przetransportowany do cytoplazmy.
Począwszy od dwóch acetylo-CoA, jeden z nich jest przekształcany w malonylo-CoA w procesie karboksylacji katalizowanej przez enzym karboksylazę acetylo-CoA (ACC), jedyny enzym regulacyjny syntezy kwasów tłuszczowych (rysunek \). Następnie, obie cząsteczki mają zastąpione części CoA przez białko nośnikowe znane jako ACP (acyl-carrier protein), tworząc acetylo-ACP i malonylo-ACP. Połączenie acylo-ACP (w tym przypadku acetylo-ACP) z malonylo-ACP powoduje rozszczepienie dodanego karboksylu i powstanie produktu pośredniego widocznego w prawym górnym rogu rysunku po lewej stronie.

Rysunek \(\PageIndex{1}): Synteza kwasów tłuszczowych
Od tego momentu reakcje chemiczne przypominają reakcje odwróconego utleniania beta. Po pierwsze, keton jest redukowany do hydroksylu przy użyciu NADPH. W przeciwieństwie do hydroksylowanego półproduktu utleniania beta, półprodukt beta jest w konfiguracji D. Następnie, woda jest usuwana z węgli 2 i 3 pośredniego hydroksylu, tworząc cząsteczkę z podwójnym wiązaniem trans. Wreszcie, podwójne wiązanie jest uwodornione, aby otrzymać nasycony półprodukt. Proces ten przebiega cyklicznie z dodawaniem kolejnych malonylo-ACP do rosnącego łańcucha, aż w końcu powstaje półprodukt o 16 karbonach (palmitoilo-CoA). W tym momencie synteza cytoplazmatyczna ustaje.
Enzymów syntezy kwasów tłuszczowych
Karboksylaza acetylo-CoA, która katalizuje syntezę malonylo-CoA, jest jedynym regulowanym enzymem w syntezie kwasów tłuszczowych. Jego regulacja obejmuje zarówno kontrolę allosteryczną, jak i modyfikację kowalencyjną. Enzym ten jest fosforylowany zarówno przez kinazę AMP, jak i kinazę białkową A. Deposforylacja jest stymulowana przez fosfatazy aktywowane przez insulinę. Deprofosforylacja aktywuje enzym i sprzyja jego łączeniu się w długi polimer, podczas gdy fosforylacja odwraca ten proces.Cytrynian działa jako aktywator allosteryczny i może również sprzyjać polimeryzacji. Palmitoyl-CoA allosterycznie go inaktywuje.
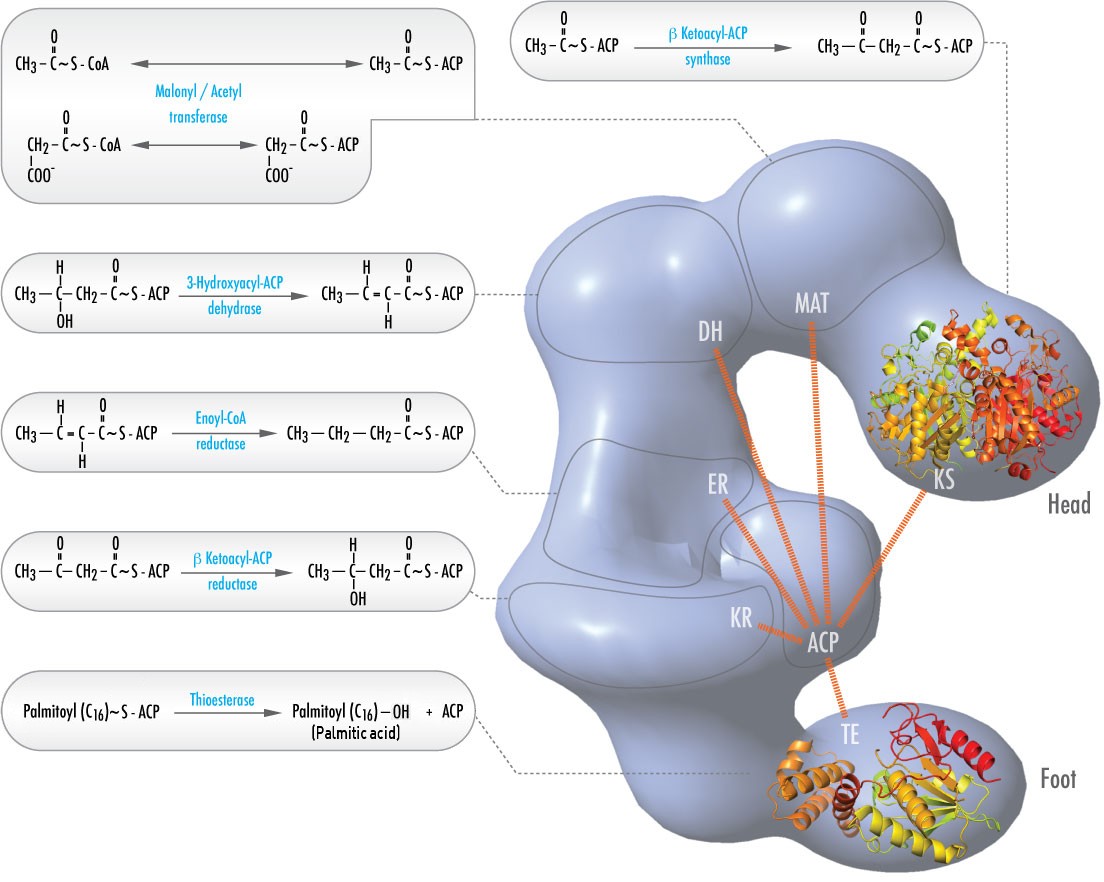
W zwierzętach, sześć różnych aktywności katalitycznych niezbędnych do pozostałych działań katalitycznych w celu pełnego wytworzenia palmitoilo-CoA jest zawartych w pojedynczym kompleksie zwanym syntazą kwasów tłuszczowych (rysunek \). Obejmują one transacylazy do zamiany CoA z ACP na acetylo-CoA i malonylo-CoA; syntazę do katalizowania dodawania jednostki dwu węglowej z trój węglowego malonylo-ACP w pierwszym etapie procesu wydłużania; reduktazę do redukcji ketonu; dehydrazę do katalizowania usuwania wody oraz reduktazę do redukcji podwójnego wiązania trans. U bakterii, te aktywności występują na oddzielnych enzymach i nie są częścią kompleksu.
Dłuższe działanie kwasów tłuszczowych
Dłuższe działanie w celu wytworzenia kwasów tłuszczowych dłuższych niż 16 karbonów zachodzi w retikulum endoplazmatycznym i jest katalizowane przez enzymy określane jako elongazy. Mitochondria również mogą wydłużać kwasy tłuszczowe, ale ich materiały wyjściowe są zazwyczaj krótsze niż 16 karbonów. Mechanizmy w obu środowiskach są podobne do tych w cytoplazmie (np. grupa malonylowa jest używana do dodania dwóch węgli), ale do produktów pośrednich przyłączany jest CoA, a nie ACP. Ponadto, podczas gdy synteza cytoplazmatyczna wykorzystuje kompleks syntazy kwasów tłuszczowych (rysunek \), enzymy w tych organellach są oddzielne i nie stanowią części kompleksu.
Desaturacja kwasów tłuszczowych
Kwasy tłuszczowe są syntetyzowane w formie nasyconej, a desaturacja następuje później. Enzymy zwane desaturazami katalizują powstawanie wiązań podwójnych cis w dojrzałych kwasach tłuszczowych. Enzymy te znajdują się w retikulum endoplazmatycznym. Zwierzęta mają ograniczoną ilość zdesaturowanych kwasów tłuszczowych, które mogą wytworzyć, ze względu na niezdolność do katalizowania reakcji poza karbony 9 i 10. Tak więc, ludzie mogą tworzyć kwas oleinowy, ale nie mogą syntetyzować kwas linolowy lub kwas linolenowy. W związku z tym te dwa muszą być dostarczane w diecie i są określane jako niezbędne kwasy tłuszczowe.
Wykonawcy
-
Dr Kevin Ahern i dr Indira Rajagopal (Oregon State University)