Lernziele
- Nennen Sie die Komponenten, die den Sympathikus und den Parasympathikus des autonomen Nervensystems erzeugen
- Erläutern Sie die Unterschiede in den Ausgangs Verbindungen innerhalb der beiden Abteilungen des autonomen Nervensystems
- Beschreiben Sie die Signalmoleküle und Rezeptorproteine, die an der Kommunikation innerhalb der beiden Abteilungen des autonomen Nervensystems beteiligt sind
Das Nervensystem kann in zwei funktionelle Teile unterteilt werden: Das somatische Nervensystem und das autonome Nervensystem. Die Hauptunterschiede zwischen den beiden Systemen zeigen sich in den Reaktionen, die sie jeweils hervorrufen. Das somatische Nervensystem bewirkt die Kontraktion der Skelettmuskulatur. Das autonome Nervensystem steuert die Herz- und glatte Muskulatur sowie das Drüsengewebe. Das somatische Nervensystem wird mit freiwilligen Reaktionen in Verbindung gebracht (obwohl viele davon ohne Bewusstsein ablaufen können, wie z. B. die Atmung), während das autonome Nervensystem mit unwillkürlichen Reaktionen in Verbindung gebracht wird, wie z. B. denjenigen, die mit der Homöostase zusammenhängen.
Das autonome Nervensystem reguliert viele der inneren Organe durch ein Gleichgewicht von zwei Aspekten oder Bereichen. In Verbindung mit dem endokrinen System ist das autonome Nervensystem maßgeblich an den homöostatischen Mechanismen im Körper beteiligt. Die beiden Bereiche des autonomen Nervensystems sind der Sympathikus und der Parasympathikus. Der Sympathikus wird mit der „Kampf-oder-Flucht“-Reaktion in Verbindung gebracht, während die parasympathische Aktivität mit dem Beinamen „Ruhe und Verdauung“ bezeichnet wird. Die Homöostase ist das Gleichgewicht zwischen den beiden Systemen. An jedem Zieleffektor bestimmt die duale Innervation die Aktivität. Zum Beispiel erhält das Herz Verbindungen sowohl vom Sympathikus als auch vom Parasympathikus. Die eine bewirkt, dass die Herzfrequenz steigt, während die andere, dass die Herzfrequenz sinkt.
Schauen Sie sich dieses Video an, um mehr über Adrenalin und die Kampf-oder-Flucht-Reaktion zu erfahren.
Wenn man von einem Adrenalinstoß spricht, kommt einem meist das Bild von Bungee-Springern oder Fallschirmspringern in den Sinn. Aber Adrenalin, auch bekannt als Epinephrin, ist eine wichtige Chemikalie bei der Koordination der Kampf-oder-Flucht-Reaktion des Körpers. In diesem Video sehen Sie einen Einblick in die Physiologie der Kampf-oder-Flucht-Reaktion, wie sie sich ein Feuerwehrmann vorstellt. Die Reaktion seines Körpers ist das Ergebnis der sympathischen Abteilung des autonomen Nervensystems, die systemweite Veränderungen verursacht, während sie sich auf extreme Reaktionen vorbereitet. Welche zwei Veränderungen bewirkt Adrenalin, um die Reaktion der Skelettmuskulatur zu unterstützen?
Sympathikus-Abteilung des autonomen Nervensystems
Um auf eine Bedrohung zu reagieren – zu kämpfen oder zu fliehen – verursacht der Sympathikus unterschiedliche Effekte, da viele verschiedene Effektororgane für einen gemeinsamen Zweck aktiviert werden. Es muss mehr Sauerstoff eingeatmet und an die Skelettmuskulatur abgegeben werden. Das Atmungs-, Herz-Kreislauf- und Muskel-Skelett-System werden gemeinsam aktiviert. itionell verhindert das Schwitzen, dass die überschüssige Wärme, die durch die Muskelkontraktion entsteht, den Körper überhitzt. Das Verdauungssystem wird heruntergefahren, damit das Blut keine Nährstoffe aufnimmt, wenn es die Skelettmuskeln mit Sauerstoff versorgen sollte. Um all diese Reaktionen zu koordinieren, verzweigen sich die Verbindungen im sympathischen System von einer begrenzten Region des Zentralnervensystems (ZNS) zu einer Vielzahl von Ganglien, die gleichzeitig zu den vielen Effektororganen projizieren. Die komplexen Strukturen, die den Ausgang des sympathischen Systems bilden, machen es möglich, dass diese disparaten Effektoren in einer koordinierten, systemischen Veränderung zusammenkommen.
Der sympathische Teil des autonomen Nervensystems beeinflusst die verschiedenen Organsysteme des Körpers durch Verbindungen, die vom thorakalen und oberen lumbalen Rückenmark ausgehen. Um diese anatomische Grundlage widerzuspiegeln, wird es als thorakolumbales System bezeichnet. Ein zentrales Neuron im Seitenhorn einer dieser Spinalregionen projiziert über die ventralen Spinalwurzeln zu Ganglien, die an die Wirbelsäule angrenzen.
Die meisten Ganglien des sympathischen Systems gehören zu einem Netzwerk von sympathischen Kettenganglien, das entlang der Wirbelsäule verläuft. Die Ganglien erscheinen als eine Reihe von Clustern von Neuronen, die durch axonale Brücken verbunden sind. Es gibt typischerweise 23 Ganglien in der Kette auf beiden Seiten der Wirbelsäule. Drei entsprechen dem zervikalen Bereich, 12 befinden sich im thorakalen Bereich, vier im lumbalen Bereich und vier im sakralen Bereich. Die zervikale und sakrale Ebene sind nicht direkt über die Spinalwurzeln mit dem Rückenmark verbunden, sondern über auf- oder absteigende Verbindungen durch die Brücken innerhalb der Kette.
Ein Diagramm, das die Verbindungen des sympathischen Systems zeigt, ist in etwa wie ein Schaltplan, der die elektrischen Verbindungen zwischen verschiedenen Gefäßen und Geräten zeigt. In Abbildung 1 sind die „Schaltkreise“ des sympathischen Systems absichtlich vereinfacht dargestellt.

Abbildung 1. Verbindungen der sympathischen Abteilung des autonomen Nervensystems. Neuronen aus dem Seitenhorn des Rückenmarks (präganglionäre Neuronen) projizieren zu den Kettenganglien auf beiden Seiten der Wirbelsäule oder zu kollateralen (prävertebralen) Ganglien, die sich anterior der Wirbelsäule in der Bauchhöhle befinden. Die Axone dieser ganglionären Neuronen (postganglionäre Fasern) projizieren dann zu Zieleffektoren im ganzen Körper.
Um die Analogie des Schaltkreises fortzusetzen, gibt es drei verschiedene Arten von „Verzweigungen“, die innerhalb des sympathischen Systems arbeiten (Abbildung 2). Der erste Typ ist der direkteste: Der sympathische Nerv projiziert zum Kettenganglion auf der gleichen Ebene wie der Zieleffektor (das Organ, Gewebe oder die Drüse, die innerviert werden soll).
Ein Beispiel für diesen Typ ist der Spinalnerv T1, der mit dem Kettenganglion T1 synaptiert, um die Luftröhre zu innervieren. Die Fasern dieses Astes werden als weiße Rami communicantes (Singular = ramus communicans) bezeichnet; sie sind myelinisiert und werden daher als weiß bezeichnet (siehe Abbildung 2a). Das Axon des zentralen Neurons (die präganglionäre Faser, dargestellt als durchgezogene Linie) synaptiert mit dem ganglionären Neuron (mit der postganglionären Faser, dargestellt als gestrichelte Linie). Dieses Neuron projiziert dann zu einem Zieleffektor – in diesem Fall die Luftröhre – über die grauen Rami communicantes, die unmyelinisierte Axone sind.
In einigen Fällen befinden sich die Zieleffektoren superior oder inferior des Spinalsegments, an dem die präganglionäre Faser austritt. Was die „Verdrahtung“ betrifft, so findet die Synapse mit dem ganglionären Neuron an den Kettenganglien superior oder inferior zum Ort des zentralen Neurons statt. Ein Beispiel hierfür ist der Spinalnerv T1, der das Auge innerviert. Der Spinalnerv zieht durch die Kette nach oben, bis er das zervikale Ganglion superior erreicht, wo er mit dem postganglionären Neuron synaptiert (siehe Abbildung 2b). Die zervikalen Ganglien werden aufgrund ihrer Lage neben den prävertebralen Ganglien in der sympathischen Kette als paravertebrale Ganglien bezeichnet.
Nicht alle Axone aus den zentralen Neuronen enden in den Kettenganglien. itionale Äste aus der ventralen Nervenwurzel setzen sich durch die Kette und weiter zu einer der kollateralen Ganglien als Nervus splanchnicus major oder Nervus splanchnicus minor fort. Zum Beispiel synaptiert der N. splanchnicus major auf der Höhe von T5 mit einem Kollateralganglion außerhalb der Kette, bevor er die Verbindung zu den postganglionären Nerven herstellt, die den Magen innervieren (siehe Abbildung 2c).
Kollateralganglien, auch prävertebrale Ganglien genannt, befinden sich anterior der Wirbelsäule und erhalten Inputs von splanchnischen Nerven sowie von zentralen sympathischen Neuronen. Sie sind mit der Steuerung von Organen in der Bauchhöhle verbunden und werden auch als Teil des enterischen Nervensystems betrachtet. Die drei Kollateralganglien sind das Ganglion celiacum, das Ganglion mesentericum superior und das Ganglion mesentericum inferior (siehe Abbildung 1). Das Wort Zöliakie leitet sich vom lateinischen Wort „coelom“ ab, das sich auf eine Körperhöhle (in diesem Fall die Bauchhöhle) bezieht, und das Wort Mesenterium bezieht sich auf das Verdauungssystem.

Abbildung 2. Sympathische Verbindungen und Kettenganglien. Das Axon eines zentralen sympathischen Neurons im Rückenmark kann auf verschiedene Weise in die Peripherie projizieren. (a) Die Faser kann zum Ganglion auf der gleichen Ebene projizieren und an einem ganglionären Neuron synapsen. (b) Ein Zweig kann zu einem höheren oder niedrigeren Ganglion in der Kette projizieren. (c) Ein Zweig kann durch den weißen Ramus communicans projizieren, aber nicht an einem ganglionären Neuron in der Kette enden. Stattdessen projiziert er durch einen der splanchnischen Nerven zu einem kollateralen Ganglion oder dem Nebennierenmark (nicht abgebildet).
Ein Axon des zentralen Neurons, das zu einem sympathischen Ganglion projiziert, wird als präganglionäre Faser oder Neuron bezeichnet und stellt den Ausgang vom ZNS zum Ganglion dar. Da die sympathischen Ganglien an die Wirbelsäule angrenzen, sind die präganglionären sympathischen Fasern relativ kurz und sie sind myelinisiert. Eine postganglionäre Faser – das Axon eines ganglionären Neurons, das zum Zieleffektor projiziert – stellt den Ausgang eines Ganglions dar, der das Organ direkt beeinflusst.
Im Vergleich zu den präganglionären Fasern sind die postganglionären sympathischen Fasern aufgrund der relativ größeren Entfernung vom Ganglion zum Zieleffektor lang. Diese Fasern sind nicht myelinisiert. (Beachten Sie, dass der Begriff „postganglionäres Neuron“ verwendet werden kann, um die Projektion von einem Ganglion zum Ziel zu beschreiben. Das Problem bei dieser Verwendung ist, dass sich der Zellkörper im Ganglion befindet, und nur die Faser postganglionär ist. Normalerweise bezieht sich der Begriff Neuron auf die gesamte Zelle.)
Eine Art von präganglionärer sympathischer Faser endet nicht in einem Ganglion. Dies sind die Axone von zentralen sympathischen Neuronen, die in das Nebennierenmark, den inneren Teil der Nebenniere, projizieren. Diese Axone werden immer noch als präganglionäre Fasern bezeichnet, aber das Ziel ist kein Ganglion. Das Nebennierenmark gibt Signalmoleküle in den Blutkreislauf ab, anstatt Axone zur Kommunikation mit Zielstrukturen zu verwenden. Die Zellen im Nebennierenmark, die von den präganglionären Fasern kontaktiert werden, heißen chromaffine Zellen. Diese Zellen sind neurosekretorische Zellen, die sich zusammen mit den sympathischen Ganglien aus der Neuralleiste entwickeln, was die Vorstellung verstärkt, dass die Drüse funktionell ein sympathisches Ganglion ist.
Die Projektionen der sympathischen Abteilung des autonomen Nervensystems gehen weit auseinander, was zu einem breiten Einfluss des Systems im gesamten Körper führt. Als Reaktion auf eine Bedrohung würde der Sympathikus die Herz- und Atemfrequenz erhöhen und bewirken, dass die Durchblutung der Skelettmuskulatur zunimmt und die Durchblutung des Verdauungssystems abnimmt. Die Schweißdrüsensekretion sollte ebenfalls als Teil einer integrierten Reaktion zunehmen.
Alle diese physiologischen Veränderungen müssen zusammen auftreten, um vor der jagenden Löwin oder dem modernen Äquivalent wegzulaufen. Diese Divergenz zeigt sich in den Verzweigungsmustern der präganglionären sympathischen Neuronen – ein einzelnes präganglionäres sympathisches Neuron kann 10-20 Ziele haben. Ein Axon, das ein zentrales Neuron des Seitenhorns im thorakolumbalen Rückenmark verlässt, passiert den weißen Ramus communicans und tritt in die sympathische Kette ein, wo es sich zu einer Vielzahl von Zielen verzweigt. Auf der Ebene des Rückenmarks, auf der die präganglionäre sympathische Faser das Rückenmark verlässt, wird ein Zweig an einem Neuron im benachbarten Kettenganglion synapsen.
Einige Zweige erstrecken sich nach oben oder unten zu einer anderen Ebene der Kettenganglien. Andere Äste durchziehen die Kettenganglien und projizieren durch einen der splanchnischen Nerven zu einem Kollateralganglion. Schließlich können einige Äste durch die splanchnischen Nerven zum Nebennierenmark projizieren. Alle diese Verzweigungen bedeuten, dass ein präganglionäres Neuron verschiedene Regionen des sympathischen Systems sehr breit beeinflussen kann, indem es auf weit verteilte Organe einwirkt.
Parasympathische Abteilung des autonomen Nervensystems
Die parasympathische Abteilung des autonomen Nervensystems ist so benannt, weil sich ihre zentralen Neuronen auf beiden Seiten der thorakolumbalen Region des Rückenmarks befinden (para- = „neben“ oder „nahe“). Der Parasympathikus kann auch als kraniosakrales System (oder Ausfluss) bezeichnet werden, weil die präganglionären Neuronen in Kernen des Hirnstamms und des Seitenhorns des sakralen Rückenmarks liegen.
Die Verbindungen oder „Schaltkreise“ der parasympathischen Abteilung ähneln dem allgemeinen Aufbau der sympathischen Abteilung mit einigen spezifischen Unterschieden (Abbildung 3). Die präganglionären Fasern aus dem kranialen Bereich verlaufen in Hirnnerven, während die präganglionären Fasern aus dem sakralen Bereich in Spinalnerven verlaufen. Die Ziele dieser Fasern sind terminale Ganglien, die sich in der Nähe – oder sogar innerhalb – des Zieleffektors befinden. Diese Ganglien werden oft als intramurale Ganglien bezeichnet, wenn sie sich innerhalb der Wände des Zielorgans befinden. Die postganglionäre Faser projiziert von den terminalen Ganglien eine kurze Strecke zum Zieleffektor oder zum spezifischen Zielgewebe innerhalb des Organs. Vergleicht man die relativen Längen der Axone im parasympathischen System, so sind die präganglionären Fasern lang und die postganglionären Fasern kurz, weil die Ganglien nahe an – und manchmal innerhalb – der Zieleffektoren liegen.
Die kraniale Komponente des parasympathischen Systems ist in bestimmten Kernen des Hirnstamms angesiedelt. Im Mittelhirn ist der Eddinger-Westphal-Kern Teil des okulomotorischen Komplexes, und die Axone dieser Neuronen wandern mit den Fasern des Nervus oculomotorius (Hirnnerv III), der die extraokulären Muskeln innerviert. Die präganglionären parasympathischen Fasern des Hirnnervs III enden im Ganglion ciliare, das sich in der hinteren Augenhöhle befindet. Die postganglionären parasympathischen Fasern projizieren dann auf die glatte Muskulatur der Iris, um die Pupillengröße zu kontrollieren. In der oberen Medulla enthalten die Speichelkerne Neuronen mit Axonen, die durch die Gesichts- und Glossopharyngeusnerven zu Ganglien projizieren, die die Speicheldrüsen kontrollieren. Die Tränenproduktion wird durch parasympathische Fasern im Nervus facialis beeinflusst, die ein Ganglion und schließlich die Tränendrüse aktivieren.
Neuronen im dorsalen Nucleus des Nervus vagus und im Nucleus ambiguus projizieren durch den Nervus vagus (Hirnnerv X) zu den terminalen Ganglien der Brust- und Bauchhöhle. Parasympathische präganglionäre Fasern beeinflussen in der Brusthöhle vor allem Herz, Bronchien und Speiseröhre und in der Bauchhöhle Magen, Leber, Pankreas, Gallenblase und Dünndarm. Die postganglionären Fasern aus den vom Nervus vagus aktivierten Ganglien sind oft in die Struktur des Organs eingebunden, wie z.B. der Plexus mesentericus der Organe des Verdauungstraktes und die intramuralen Ganglien.

Abbildung 3. Verbindungen der parasympathischen Abteilung des autonomen Nervensystems Neuronen aus Hirnstammkernen oder aus dem Seitenhorn des sakralen Rückenmarks projizieren zu terminalen Ganglien in der Nähe oder innerhalb der verschiedenen Organe des Körpers. Axone von diesen Ganglienneuronen projizieren dann die kurze Strecke zu diesen Zieleffektoren.
Chemische Signalübertragung im autonomen Nervensystem
Wo sich ein autonomes Neuron mit einem Ziel verbindet, entsteht eine Synapse. Das elektrische Signal des Aktionspotentials bewirkt die Freisetzung eines Signalmoleküls, das an Rezeptorproteine auf der Zielzelle bindet. Synapsen des autonomen Systems werden entweder als cholinergisch klassifiziert, was bedeutet, dass Acetylcholin (ACh) freigesetzt wird, oder adrenergisch, was bedeutet, dass Noradrenalin freigesetzt wird. Die Begriffe cholinergisch und adrenergisch beziehen sich nicht nur auf das Signalmolekül, das freigesetzt wird, sondern auch auf die Klasse der Rezeptoren, die jeweils gebunden werden.
Das cholinerge System umfasst zwei Klassen von Rezeptoren: den Nikotinrezeptor und den Muskarinrezeptor. Beide Rezeptortypen binden an ACh und bewirken Veränderungen in der Zielzelle. Der Nikotinrezeptor ist ein ligandengesteuerter Kationenkanal und der Muskarinrezeptor ist ein G-Protein-gekoppelter Rezeptor. Die Rezeptoren werden nach anderen Molekülen benannt, die an sie binden, und unterscheiden sich dadurch. Während Nikotin an den nicotinischen Rezeptor und Muscarin an den muscarinischen Rezeptor bindet, gibt es keine Kreuzreaktivität zwischen den Rezeptoren. Die Situation ist ähnlich wie bei Schlössern und Schlüsseln.
Stellen Sie sich zwei Schlösser vor – eines für ein Klassenzimmer und das andere für ein Büro -, die mit zwei verschiedenen Schlüsseln geöffnet werden. Der Schlüssel für das Klassenzimmer öffnet nicht die Tür des Büros und der Schlüssel für das Büro öffnet nicht die Tür des Klassenzimmers. Dies ist ähnlich wie die Spezifität von Nikotin und Muscarin für ihre Rezeptoren. Ein Hauptschlüssel kann jedoch mehrere Schlösser öffnen, z. B. ein Hauptschlüssel für die Biologieabteilung, der sowohl die Klassenzimmer- als auch die Bürotür öffnet. Dies ist ähnlich wie bei ACh, das an beide Arten von Rezeptoren bindet. Die Moleküle, die diese Rezeptoren definieren, sind nicht entscheidend – sie sind einfach Werkzeuge für die Forscher, die sie im Labor verwenden. Diese Moleküle sind exogen, was bedeutet, dass sie außerhalb des menschlichen Körpers hergestellt werden, so dass ein Forscher sie ohne störende endogene Ergebnisse (Ergebnisse, die durch die im Körper produzierten Moleküle verursacht werden) verwenden kann.
Das adrenerge System verfügt ebenfalls über zwei Arten von Rezeptoren, den alpha (α)-adrenergen Rezeptor und den beta (β)-adrenergen Rezeptor. Im Gegensatz zu cholinergen Rezeptoren werden diese Rezeptortypen nicht danach klassifiziert, welche Medikamente an sie binden können. Sie alle sind G-Protein-gekoppelte Rezeptoren. Es gibt drei Typen von α-adrenergen Rezeptoren, die als α1, α2 und α3 bezeichnet werden, und es gibt zwei Typen von β-adrenergen Rezeptoren, die als β1 und β2 bezeichnet werden. Ein zusätzlicher Aspekt des adrenergen Systems ist, dass es ein zweites Signalmolekül gibt, das Epinephrin genannt wird. Der chemische Unterschied zwischen Norepinephrin und Epinephrin ist das Vorhandensein einer Methylgruppe (CH3) im Epinephrin. Die Vorsilbe „nor-“ bezieht sich eigentlich auf diesen chemischen Unterschied, bei dem eine Methylgruppe fehlt.
Der Begriff adrenerge soll an das Wort Adrenalin erinnern, das mit der zu Beginn des Kapitels beschriebenen Kampf-oder-Flucht-Reaktion in Verbindung gebracht wird. Adrenalin und Epinephrin sind zwei Namen für dasselbe Molekül. Die Nebenniere (lateinisch ad- = „oben auf“; renal = „Niere“) sondert Adrenalin ab. Die Endung „-ine“ bezieht sich darauf, dass die Chemikalie von der Nebenniere abgeleitet bzw. extrahiert wird. Eine ähnliche Konstruktion aus dem Griechischen statt aus dem Lateinischen ergibt das Wort Epinephrin (epi- = „oben“; nephr- = „Niere“). Im wissenschaftlichen Sprachgebrauch wird in den Vereinigten Staaten Epinephrin bevorzugt, während in Großbritannien Adrenalin bevorzugt wird, da „Adrenalin“ in den Vereinigten Staaten einst ein registrierter, geschützter Medikamentenname war. Obwohl das Medikament nicht mehr verkauft wird, bleibt die Konvention bestehen, dieses Molekül mit den beiden unterschiedlichen Namen zu bezeichnen. In ähnlicher Weise sind Noradrenalin und Noradrenalin zwei Namen für dasselbe Molekül.
Nachdem man das cholinerge und adrenerge System verstanden hat, ist ihre Rolle im autonomen System relativ einfach zu verstehen. Alle präganglionären Fasern, sowohl des Sympathikus als auch des Parasympathikus, setzen ACh frei. Alle ganglionären Neuronen – die Ziele dieser präganglionären Fasern – haben nicotinische Rezeptoren in ihren Zellmembranen. Der Nikotinrezeptor ist ein ligandengesteuerter Kationenkanal, der zu einer Depolarisation der postsynaptischen Membran führt. Die postganglionären parasympathischen Fasern setzen ebenfalls ACh frei, aber die Rezeptoren auf ihren Zielen sind Muscarinrezeptoren, die G-Protein-gekoppelte Rezeptoren sind und nicht ausschließlich eine Depolarisation der postsynaptischen Membran bewirken. Postganglionäre sympathische Fasern setzen Noradrenalin frei, mit Ausnahme der Fasern, die zu den Schweißdrüsen und zu den mit den Skelettmuskeln verbundenen Blutgefäßen projizieren, die ACh freisetzen (Tabelle 1).
| Tabelle 1 | ||
|---|---|---|
| Sympathikus | Parasympathikus | |
| Preganglionär | Acetylcholin>Nikotinrezeptor | Acetylcholin> Nikotinrezeptor |
| Postganglionär | Norepinephrin > a- oder B-adrenerge Rezeptoren Acetylcholin > muskarinischer Rezeptor (nur mit Schweißdrüsen und den Blutgefäßen der Skelettmuskulatur verbunden |
Acetylcholin > muskarinischer Rezeptor |
Signalmoleküle können zu zwei großen Gruppen gehören. Neurotransmitter werden an den Synapsen freigesetzt, während Hormone in den Blutkreislauf abgegeben werden. Dies sind vereinfachte Definitionen, aber sie können helfen, diesen Punkt zu klären. Acetylcholin kann als Neurotransmitter betrachtet werden, weil es von Axonen an Synapsen freigesetzt wird. Das adrenerge System stellt jedoch eine Herausforderung dar. Postganglionäre sympathische Fasern setzen Noradrenalin frei, das als Neurotransmitter betrachtet werden kann. Aber das Nebennierenmark setzt Epinephrin und Norepinephrin in den Kreislauf frei, so dass sie als Hormone betrachtet werden sollten.
Was hier als Synapsen bezeichnet wird, entspricht nicht unbedingt der strengsten Definition von Synapse. Einige Quellen bezeichnen die Verbindung zwischen einer postganglionären Faser und einem Zieleffektor als Neuroeffector Junctions; Neurotransmitter, wie oben definiert, würden als Neuromodulatoren bezeichnet werden. Die Struktur der postganglionären Verbindungen ist nicht der typische synaptische Endknollen, der an der neuromuskulären Verbindung zu finden ist, sondern es sind Ketten von Schwellungen entlang der Länge einer postganglionären Faser, die Varicosity genannt werden (Abbildung 4).
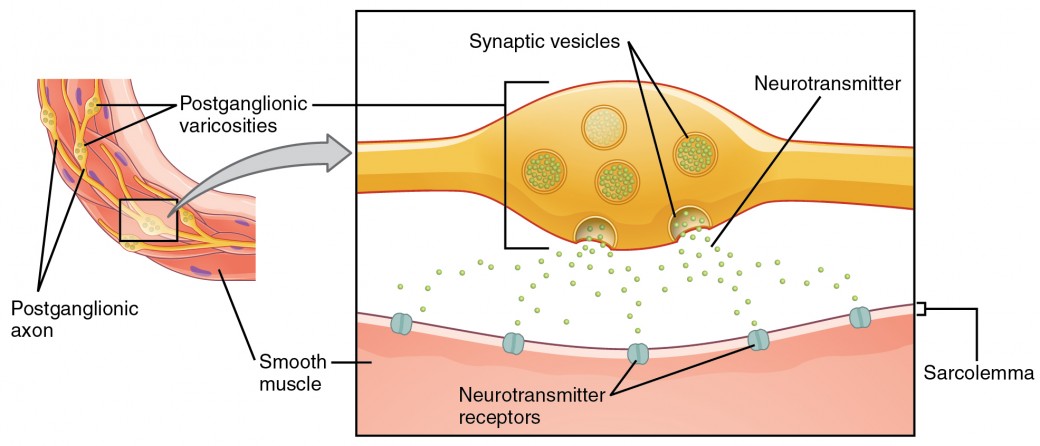
Abbildung 4. Autonome Varizen. Die Verbindung zwischen autonomen Fasern und Zieleffektoren entspricht nicht der typischen Synapse, wie zum Beispiel der neuromuskulären Verbindung. Anstelle eines synaptischen Endzwiebels wird ein Neurotransmitter aus Schwellungen entlang der Länge einer Faser freigesetzt, die ein ausgedehntes Netzwerk von Verbindungen im Zieleffektor bildet.
Alltägliche Verbindungen: Fight or Flight? Was ist mit Schreck und Erstarren?
Die ursprüngliche Verwendung des Beinamens „Kampf oder Flucht“ stammt von einem Wissenschaftler namens Walter Cannon, der 1915 in Harvard arbeitete. Das Konzept der Homöostase und der Funktionsweise des Sympathikus war bereits im vorigen Jahrhundert in Frankreich eingeführt worden. Cannon erweiterte die Idee und führte die Vorstellung ein, dass ein Tier auf eine Bedrohung reagiert, indem es sich darauf vorbereitet, zu stehen und zu kämpfen oder wegzulaufen. Die Natur dieser Reaktion wurde in einem Buch über die Physiologie von Schmerz, Hunger, Angst und Wut gründlich erklärt.
Wenn Studenten etwas über das sympathische System und die Kampf-oder-Flucht-Reaktion lernen, halten sie oft inne und fragen sich nach anderen Reaktionen. Wenn Sie mit einer Löwin konfrontiert wären, die auf Sie zu rennt, wie zu Beginn dieses Kapitels abgebildet, würden Sie weglaufen oder würden Sie standhaft bleiben? Manche Menschen würden sagen, sie würden erstarren und nicht wissen, was sie tun sollen. Gibt es also nicht wirklich mehr, was das autonome System tut, als Kampf, Flucht, Ruhe oder Verdauung. Was ist mit Angst und Lähmung im Angesicht einer Bedrohung?
Das gängige Schlagwort von „Kampf oder Flucht“ wird erweitert zu „Kampf, Flucht oder Angst“ oder sogar „Kampf, Flucht, Angst oder Erstarren.“ Cannons ursprünglicher Beitrag war eine eingängige Phrase, um etwas von dem auszudrücken, was das Nervensystem als Reaktion auf eine Bedrohung tut, aber sie ist unvollständig. Das sympathische System ist für die physiologischen Reaktionen auf emotionale Zustände verantwortlich. Der Name „Sympathikus“ kann das ausdrücken (sym- = „zusammen“; -pathos = „Schmerz“, „Leiden“ oder „Emotion“).
Schauen Sie sich dieses Video an, um mehr über das Nervensystem zu erfahren.
Wie in diesem Video beschrieben, hat das Nervensystem einen Weg, mit Bedrohungen und Stress umzugehen, der von der bewussten Kontrolle des somatischen Nervensystems getrennt ist. Das System stammt aus einer Zeit, in der es bei Bedrohungen um das Überleben ging, aber in der modernen Zeit werden diese Reaktionen zu einem Teil von Stress und Angst. Dieses Video beschreibt, dass das autonome System nur ein Teil der Reaktion auf Bedrohungen oder Stressoren ist. Welches andere Organsystem ist beteiligt, und welcher Teil des Gehirns koordiniert die beiden Systeme für die gesamte Reaktion, einschließlich Adrenalin und Cortisol?