Beurteilung | Biopsychologie | Vergleichend |Kognitiv | Entwicklung | Sprache | Individuelle Unterschiede |Persönlichkeit | Philosophie | Soziales |
Methoden | Statistik |Klinische | Pädagogische | Industrielle |Berufliche Gegenstände |Weltpsychologie |
Biologisch:Verhaltensgenetik – Evolutionspsychologie – Neuroanatomie – Neurochemie – Neuroendokrinologie -Neurowissenschaft – Psychoneuroimmunologie – Physiologische Psychologie – Psychopharmakologie(Index, Gliederung)
| Rod Zelle | |
|---|---|
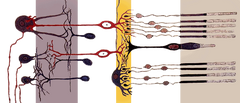
|
|
| Ort | Retina |
| Funktion | Schwachlicht-Photorezeptor |
| Morphologie | Stäbchenform |
| Presynaptische Verbindungen | Keine |
| Postsynaptische Verbindungen | Bipolarzellen und Horizontalzellen |
Stäbchenzellen, oder Stäbchen, sind Photorezeptorzellen in der Netzhaut des Auges, die bei weniger intensivem Licht funktionieren können als die andere Art von Photorezeptoren, die Zapfenzellen. Da sie lichtempfindlicher sind, sind die Stäbchen für das Nachtsehen verantwortlich. Benannt nach ihrer zylindrischen Form, sind Stäbchen an den äußeren Rändern der Netzhaut konzentriert und werden für das periphere Sehen verwendet. Es gibt etwa 120 Millionen Stäbchenzellen in der menschlichen Netzhaut.
Eine Stäbchenzelle ist empfindlich genug, um auf ein einzelnes Lichtphoton zu reagieren, und ist etwa 100 Mal empfindlicher für ein einzelnes Photon als Zapfen. Da Stäbchen weniger Licht benötigen, um zu funktionieren als Zapfen, sind sie daher die primäre Quelle für visuelle Informationen bei Nacht (skotopisches Sehen). Zapfenzellen hingegen benötigen Dutzende bis Hunderte von Photonen, um aktiviert zu werden. Außerdem konvergieren mehrere Stäbchenzellen auf ein einziges Interneuron, das die Signale sammelt und verstärkt. Diese Konvergenz geht jedoch zu Lasten der Sehschärfe (oder der Bildauflösung), da die zusammengefassten Informationen von mehreren Zellen weniger deutlich sind, als wenn das visuelle System Informationen von jeder einzelnen Stäbchenzelle erhalten würde. Die Konvergenz der Stäbchenzellen macht auch das periphere Sehen sehr empfindlich für Bewegungen und ist verantwortlich für das Phänomen, dass ein Mensch etwas Unscharfes aus dem Augenwinkel sieht.
Stäbchen sind etwas schmaler als Zapfen, haben aber die gleiche strukturelle Basis. Das Pigment befindet sich auf der Außenseite und liegt auf dem Pigmentepithel. An diesem Ende befinden sich viele gestapelte Scheiben, wahrscheinlich durch die Einfaltung der diesen Abschnitt umgebenden Grenzmembran, wodurch eine größere Fläche für das Sehpigment entsteht und die Effizienz der Lichtabsorption erhöht wird. Da sie nur eine Art von lichtempfindlichem Pigment haben, anstatt der drei Arten, die menschliche Zapfenzellen haben, spielen Stäbchen, wenn überhaupt, nur eine geringe Rolle beim Farbensehen.
Stäbchenzellen reagieren auch langsamer auf Licht als Zapfen, so dass Reize, die sie empfangen, über etwa 100 Millisekunden addiert werden. Das macht die Stäbchen zwar empfindlicher für kleinere Lichtmengen, bedeutet aber auch, dass ihre Fähigkeit, zeitliche Veränderungen, wie z. B. schnell wechselnde Bilder, wahrzunehmen, weniger genau ist als die der Zapfen.
Experimente von George Wald und anderen zeigten, dass Stäbchen am empfindlichsten auf Lichtwellenlängen um 498 nm (grün-blau) reagieren und völlig unempfindlich für Wellenlängen über etwa 640 nm (rot) sind. Diese Tatsache ist verantwortlich für den Purkinje-Effekt, bei dem in der Dämmerung, wenn die Stäbchen als die für das Sehen zuständigen Zellen übernehmen, blaue Farben im Vergleich zu roten intensiver erscheinen.
Wie die Zapfen haben auch die Stäbchenzellen ein synaptisches Terminal, ein inneres Segment und ein äußeres Segment. Das synaptische Terminal bildet eine Synapse mit einem anderen Neuron, zum Beispiel einer Bipolarzelle. Das innere und das äußere Segment sind durch ein Cilium miteinander verbunden. Das innere Segment enthält Organellen und den Zellkern, während das Stäbchenaußensegment (abgekürzt ROS), das zum Augenhintergrund gerichtet ist, die lichtabsorbierenden Materialien enthält.
Reaktion auf Licht
Anatomie einer Stäbchenzelle
Die Aktivierung eines einzelnen Moleküls von Rhodopsin, dem lichtempfindlichen Pigment in Stäbchen, kann zu einer großen Reaktion in der Zelle führen, da das Signal verstärkt wird. Einmal aktiviert, kann Rhodopsin Hunderte von Transducin-Molekülen aktivieren, von denen jedes wiederum ein Phosphodiesterase-Molekül aktiviert, das über tausend cGMP-Moleküle pro Sekunde abbauen kann. So können Stäbchen eine große Reaktion auf eine kleine Lichtmenge haben.
Da die Netzhautkomponente von Rhodopsin von Vitamin A abgeleitet wird, führt ein Mangel an Vitamin A zu einem Defizit an dem von den Stäbchenzellen benötigten Pigment. Infolgedessen können weniger Stäbchenzellen bei Dunkelheit ausreichend reagieren, und da die Zapfenzellen für das Sehen in der Dunkelheit schlecht angepasst sind, kann es zur Erblindung kommen. Dies ist die Nachtblindheit.
Rückkehr in den Ruhezustand
Die Stäbchen nutzen drei inhibitorische Mechanismen (negative Rückkopplungsmechanismen), um eine schnelle Rückkehr in den Ruhezustand nach einem Lichtblitz zu ermöglichen.
Zum einen gibt es eine Rhodopsin-Kinase (RK), die den zytosolischen Schwanz des aktivierten Rhodopsins an mehreren Serinen phosphorylieren würde, wodurch die Aktivierung von Transducin teilweise gehemmt wird. Außerdem bindet dann ein inhibitorisches Protein – das Arrestin – an die phosphorylierten Rhodopsine, um die Aktivität des Rhodopsins weiter zu hemmen.
Während Arrestin das Rhodopsin abschaltet, treibt ein RGS-Protein (das als GTPase-aktivierendes Protein (GAPs) fungiert) das Transducin (G-Protein) in einen „Aus“-Zustand, indem es die Rate der Hydrolyse des gebundenen GTP zu GDP erhöht.
Da die cGMP-sensitiven Kanäle nicht nur den Einstrom von Natrium-Ionen, sondern auch von Kalzium-Ionen zulassen, werden mit der Abnahme der cGMP-Konzentration die cGMP-sensitiven Kanäle geschlossen und damit der normale Einstrom von Kalzium-Ionen reduziert. Die Abnahme der Kalziumionenkonzentration stimuliert die kalziumionensensitiven Proteine, die dann die Guanylylzyklase aktivieren würden, um das cGMP wieder aufzufüllen, wodurch die ursprüngliche Konzentration schnell wiederhergestellt wird. Die Wiederherstellung öffnet die cGMP-empfindlichen Kanäle und bewirkt eine Depolarisation der Plasmamembran.
Desensibilisierung
Wenn die Stäbchen über einen längeren Zeitraum einer hohen Photonenkonzentration ausgesetzt sind, werden sie desensibilisiert (angepasst) an die Umgebung.
Da Rhodopsin durch die Rhodopsin-Kinase (ein Mitglied der GPCR-Kinasen (GRKs)) phosphoryliert wird, bindet es mit hoher Affinität an das Arrestin. Das gebundene Arrestin kann auf mindestens zwei Arten zum Desensibilisierungsprozess beitragen. Erstens verhindert es die Interaktion zwischen dem G-Protein und dem aktivierten Rezeptor. Zweitens dient es als Adaptorprotein, um den Rezeptor an die Clathrin-abhängige Endozytose-Maschinerie heranzuführen (um rezeptorvermittelte Endozytose zu induzieren).
Tabelle
Vergleich von Stäbchen- und Zapfenzellen, aus Kandel et al. (2000).
| Stäbchen | Zapfen |
|---|---|
| für Nachtsehen | für Tagessehen | sehr lichtempfindlich; empfindlich für Streulicht | nicht sehr lichtempfindlich; empfindlich nur für direktes Licht |
| Verlust verursacht Nachtblindheit | Verlust verursacht Rechtsblindheit |
| niedrige Sehschärfe | hohe Sehschärfe; bessere räumliche Auflösung |
| nicht in der Fovea vorhanden | konzentriert in der Fovea |
| langsame Reaktion, auf Licht, Reize werden mit der Zeit hinzugefügt | schnelle Reaktion auf Licht, können schnellere Veränderungen von Reizen wahrnehmen |
| haben mehr Pigment als Zapfen, können daher weniger Licht erkennen | haben weniger Pigment als Stäbchen, benötigen mehr Licht, um Bilder zu erkennen |
| Stapel von membraneingeschlossene Scheiben sind nicht mit der Zellmembran verbunden | Scheiben sind mit der äußeren Membran verbunden |
| 20-mal mehr Stäbchen als Zapfen in der Netzhaut | |
| ein Typ lichtempfindliches Pigment | drei Arten von lichtempfindlichen Pigmenten beim Menschen |
| vermitteln achromatisches Sehen | vermitteln Farbsehen |
Referenz
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4. Aufl., S.507-513. McGraw-Hill, New York.
- Human Physiology and Mechanisms of Disease von Arthur C. Guyton (1992) S.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Molecular Biology of The Cell, 5. Aufl., S.919-921. Garland Science. Zitierfehler: Invalid
<ref>tag; name „Alberts“ defined multiple times with different content
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4. Aufl., S.507-513. McGraw-Hill, New York.