La síntesis de ácidos grasos ocurre en el citoplasma y retículo endoplásmico de la célula y es químicamente similar al proceso de beta-oxidación, pero con un par de diferencias clave. La primera de ellas se produce en la preparación de los sustratos para las reacciones que hacen crecer el ácido graso. El transporte de acetil-CoA desde la mitocondria se produce cuando empieza a acumularse. Dos moléculas pueden desempeñar un papel en su traslado al citoplasma: el citrato y la acetilcarnitina. La unión del oxaloacetato con el acetil-CoA en la mitocondria crea citrato, que se desplaza a través de la membrana, seguido de la acción de la citrato liasa en el citoplasma de la célula para liberar acetil-CoA y oxaloacetato. Además, cuando el acetil-CoA libre se acumula en la mitocondria, puede combinarse con la carnitina y ser transportado hacia el citoplasma.
Comenzando con dos acetil-CoA, uno se convierte en malonil-CoA por carboxilación catalizada por la enzima acetil-CoA carboxilasa (ACC), la única enzima reguladora de la síntesis de ácidos grasos (Figura \N(\PageIndex{1})). A continuación, ambas moléculas tienen sus porciones de CoA sustituidas por una proteína transportadora conocida como ACP (proteína transportadora de acil) para formar acetil-ACP y malonil-ACP. La unión de un acil-ACP graso (en este caso, acetil-ACP) con el malonil-ACP escinde el carboxilo que se añadió y crea el intermedio de la parte superior derecha en la figura de la izquierda.

Figura (\PageIndex{1}): Síntesis de ácidos grasos
A partir de este punto, las reacciones químicas se asemejan a las de la beta oxidación invertida. En primer lugar, la cetona se reduce a un hidroxilo utilizando NADPH. En contraste con el intermedio hidroxilado de la beta oxidación, el intermedio beta aquí está en la configuración D. A continuación, se elimina el agua de los carbonos 2 y 3 del intermedio hidroxilo para producir una molécula de doble enlace trans. Por último, el doble enlace se hidrogena para producir un intermedio saturado. El proceso se repite con la adición de otro malonil-ACP a la cadena creciente hasta que finalmente se produce un intermediario con 16 carbonos (palmitoil-CoA). En este punto, la síntesis citoplasmática cesa.
Enzimas de la síntesis de ácidos grasos
La acetil-CoA carboxilasa, que cataliza la síntesis de malonil-CoA, es la única enzima regulada en la síntesis de ácidos grasos. Su regulación implica tanto el control alostérico como la modificación covalente. Se sabe que la enzima es fosforilada tanto por la AMP quinasa como por la proteína quinasa A. La desfosforilación es estimulada por las fosfatasas activadas por la unión de la insulina. La desfosforilación activa la enzima y favorece su ensamblaje en un polímero largo, mientras que la fosforilación invierte el proceso.El citrato actúa como activador alostérico y también puede favorecer la polimerización. El palmitoil-CoA la inactiva alostéricamente.
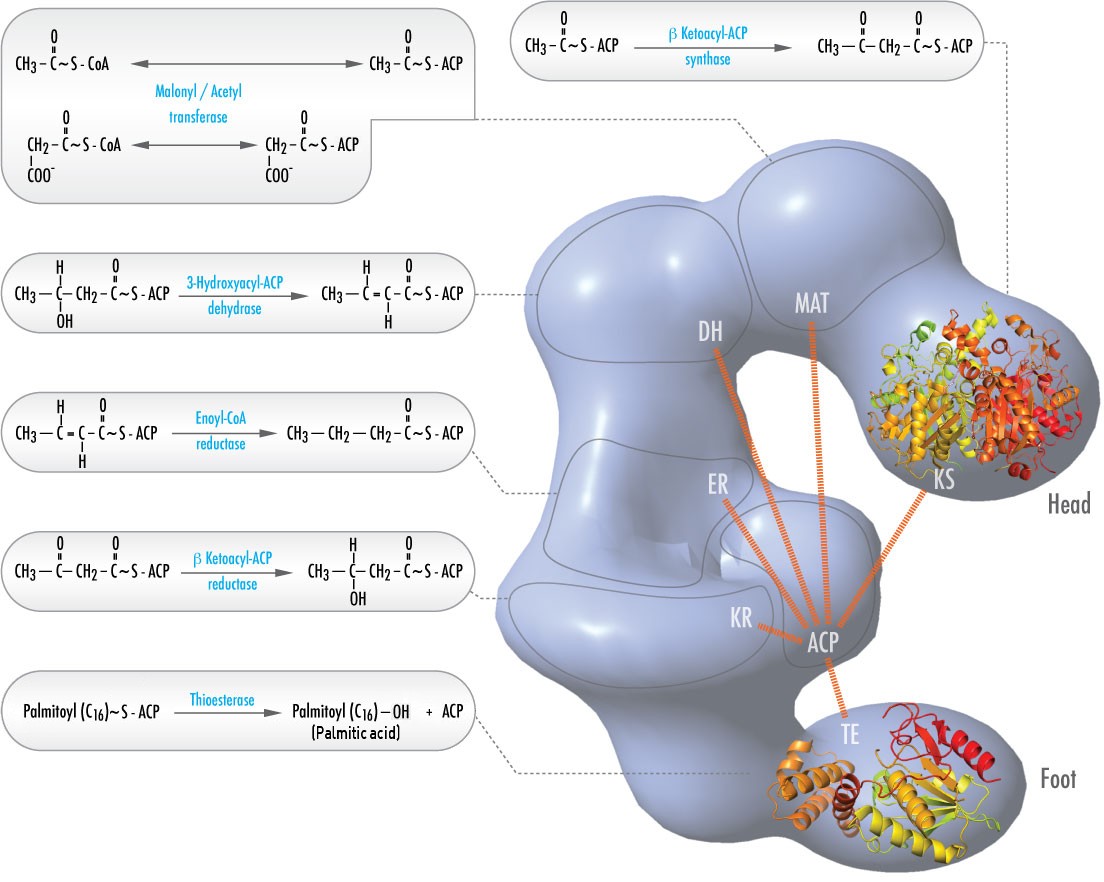
En los animales, seis actividades catalíticas diferentes necesarias para el resto de acciones catalíticas para fabricar completamente palmitoil-CoA están contenidas en un único complejo llamado Ácido Graso Sintasa (Figura \(\PageIndex{2}\)). Entre ellas se encuentran las transacilasas para intercambiar CoA con ACP en el acetil-CoA y el malonil-CoA; una sintasa para catalizar la adición de la unidad de dos carbonos del malonil-ACP de tres carbonos en el primer paso del proceso de elongación; una reductasa para reducir la cetona; una deshidrasa para catalizar la eliminación del agua, y una reductasa para reducir el doble enlace trans. En las bacterias, estas actividades se encuentran en enzimas separadas y no forman parte de un complejo.
Elongación de ácidos grasos
La elongación para hacer ácidos grasos de más de 16 carbonos ocurre en el retículo endoplásmico y es catalizada por enzimas descritas como elongasas. Las mitocondrias también pueden alargar los ácidos grasos, pero sus materiales de partida son generalmente más cortos de 16 carbonos. Los mecanismos en ambos entornos son similares a los del citoplasma (se utiliza un grupo malonilo para añadir dos carbonos, por ejemplo), pero a los intermedios se les une CoA, no ACP. Además, mientras que la síntesis citoplasmática emplea el complejo de la sintasa de ácidos grasos (Figura \(\PageIndex{2})), las enzimas en estos orgánulos son separables y no forman parte de un complejo.
Desaturación de ácidos grasos
Los ácidos grasos se sintetizan en la forma saturada y la desaturación se produce posteriormente. Unas enzimas llamadas desaturasas catalizan la formación de dobles enlaces cis en los ácidos grasos maduros. Estas enzimas se encuentran en el retículo endoplásmico. Los animales están limitados en cuanto a los ácidos grasos desaturados que pueden producir, debido a la incapacidad de catalizar las reacciones más allá de los carbonos 9 y 10. Así, los seres humanos pueden fabricar ácido oleico, pero no pueden sintetizar ácido linoleico ni ácido linolénico. En consecuencia, estos dos deben ser suministrados en la dieta y se denominan ácidos grasos esenciales.
Contribuidores
-
Dr. Kevin Ahern y Dra. Indira Rajagopal (Universidad Estatal de Oregón)