Objetivos de aprendizaje
- Nombrar los componentes que generan las respuestas simpática y parasimpática del sistema nervioso autónomo
- Explicar las diferencias en las conexiones de salida conexiones dentro de las dos divisiones del sistema nervioso autónomo
- Describa las moléculas de señalización y las proteínas receptoras implicadas en la comunicación dentro de las dos divisiones del sistema nervioso autónomo
El sistema nervioso puede dividirse en dos partes funcionales: el sistema nervioso somático y el sistema nervioso autónomo. Las principales diferencias entre ambos sistemas son evidentes en las respuestas que cada uno produce. El sistema nervioso somático provoca la contracción de los músculos esqueléticos. El sistema nervioso autónomo controla los músculos cardíacos y lisos, así como el tejido glandular. El sistema nervioso somático se asocia con las respuestas voluntarias (aunque muchas pueden ocurrir sin conciencia, como la respiración), y el sistema nervioso autónomo se asocia con las respuestas involuntarias, como las relacionadas con la homeostasis.
El sistema nervioso autónomo regula muchos de los órganos internos a través de un equilibrio de dos aspectos, o divisiones. En relación con el sistema endocrino, el sistema nervioso autónomo es fundamental en los mecanismos homeostáticos del cuerpo. Las dos divisiones del sistema nervioso autónomo son la división simpática y la división parasimpática. El sistema simpático se asocia con la respuesta de lucha o huida, y la actividad parasimpática se conoce con el epíteto de descanso y digestión. La homeostasis es el equilibrio entre los dos sistemas. En cada efector objetivo, la inervación dual determina la actividad. Por ejemplo, el corazón recibe conexiones de las divisiones simpática y parasimpática. Una de ellas hace que aumente la frecuencia cardíaca, mientras que la otra hace que disminuya.
Cuando se dice que alguien tiene un subidón de adrenalina, suele venir a la mente la imagen de los puenteros o los paracaidistas. Pero la adrenalina, también conocida como epinefrina, es una sustancia química importante para coordinar la respuesta de lucha o huida del cuerpo. En este vídeo, podrá ver la fisiología de la respuesta de lucha o huida, tal y como la concibe un bombero. La reacción de su cuerpo es el resultado de la división simpática del sistema nervioso autónomo que provoca cambios en todo el sistema mientras se prepara para respuestas extremas. ¿Qué dos cambios provoca la adrenalina para ayudar a la respuesta del músculo esquelético?
División simpática del sistema nervioso autónomo
Para responder a una amenaza -luchar o huir- el sistema simpático provoca efectos divergentes ya que muchos órganos efectores diferentes se activan juntos con un propósito común. Es necesario inhalar más oxígeno y entregarlo al músculo esquelético. Los sistemas respiratorio, cardiovascular y musculoesquelético se activan conjuntamente. ionalmente, la sudoración evita que el exceso de calor procedente de la contracción muscular provoque un sobrecalentamiento del cuerpo. El sistema digestivo se apaga para que la sangre no absorba nutrientes cuando debería llevar oxígeno a los músculos esqueléticos. Para coordinar todas estas respuestas, las conexiones del sistema simpático divergen desde una región limitada del sistema nervioso central (SNC) hasta un amplio conjunto de ganglios que se proyectan simultáneamente a los numerosos órganos efectores. El complejo conjunto de estructuras que componen la salida del sistema simpático hace posible que estos efectores dispares se unan en un cambio coordinado y sistémico.
La división simpática del sistema nervioso autónomo influye en los diversos sistemas de órganos del cuerpo a través de conexiones que surgen de la médula espinal torácica y lumbar superior. Se denomina sistema toracolumbar para reflejar esta base anatómica. Una neurona central en el cuerno lateral de cualquiera de estas regiones espinales se proyecta a los ganglios adyacentes a la columna vertebral a través de las raíces espinales ventrales.
La mayoría de los ganglios del sistema simpático pertenecen a una red de ganglios de la cadena simpática que corre a lo largo de la columna vertebral. Los ganglios aparecen como una serie de grupos de neuronas unidas por puentes axonales. Normalmente hay 23 ganglios en la cadena a cada lado de la columna vertebral. Tres corresponden a la región cervical, 12 a la torácica, cuatro a la lumbar y cuatro a la sacra. Los niveles cervical y sacro no están conectados a la médula espinal directamente a través de las raíces espinales, sino a través de conexiones ascendentes o descendentes a través de los puentes dentro de la cadena.
Un diagrama que muestra las conexiones del sistema simpático es algo así como un diagrama de circuito que muestra las conexiones eléctricas entre diferentes receptáculos y dispositivos. En la Figura 1, los «circuitos» del sistema simpático están intencionadamente simplificados.

Figura 1. Conexiones de la división simpática del sistema nervioso autónomo. Las neuronas del cuerno lateral de la médula espinal (neuronas preganglionares) se proyectan a los ganglios de la cadena a ambos lados de la columna vertebral o a los ganglios colaterales (prevertebrales) que están por delante de la columna vertebral en la cavidad abdominal. Los axones de estas neuronas ganglionares (fibras postganglionares) se proyectan entonces a los efectores diana de todo el cuerpo.
Un ejemplo de este tipo es el nervio espinal T1 que hace sinapsis con el ganglio de la cadena T1 para inervar la tráquea. Las fibras de esta rama se denominan rami comunicantes blancas (singular = ramus communicans); están mielinizadas y por ello se denominan blancas (véase la figura 2a). El axón de la neurona central (la fibra preganglionar mostrada como línea sólida) hace sinapsis con la neurona ganglionar (con la fibra postganglionar mostrada como línea discontinua). Esta neurona se proyecta entonces a un efector de destino -en este caso, la tráquea- a través de rami comunicantes grises, que son axones no mielinizados.
En algunos casos, los efectores de destino se sitúan por encima o por debajo del segmento espinal en el que emerge la fibra preganglionar. Con respecto al «cableado» implicado, la sinapsis con la neurona ganglionar se produce en los ganglios de la cadena superior o inferior a la ubicación de la neurona central. Un ejemplo de ello es el nervio espinal T1 que inerva el ojo. El nervio espinal asciende por la cadena hasta llegar al ganglio cervical superior, donde hace sinapsis con la neurona postganglionar (véase la figura 2b). Los ganglios cervicales se denominan ganglios paravertebrales, dada su ubicación adyacente a los ganglios prevertebrales en la cadena simpática.
No todos los axones de las neuronas centrales terminan en los ganglios de la cadena. Las ramas tradicionales de la raíz nerviosa ventral continúan a través de la cadena y hacia uno de los ganglios colaterales como el nervio esplácnico mayor o el nervio esplácnico menor. Por ejemplo, el nervio esplácnico mayor a nivel de T5 hace sinapsis con un ganglio colateral fuera de la cadena antes de hacer la conexión con los nervios postganglionares que inervan el estómago (ver Figura 2c).
Los ganglios colaterales, también llamados ganglios prevertebrales, están situados por delante de la columna vertebral y reciben entradas de los nervios esplácnicos así como de las neuronas simpáticas centrales. Están asociados al control de los órganos de la cavidad abdominal y también se consideran parte del sistema nervioso entérico. Los tres ganglios colaterales son el ganglio celíaco, el ganglio mesentérico superior y el ganglio mesentérico inferior (véase la figura 1). La palabra celíaco deriva de la palabra latina «coelom», que se refiere a una cavidad corporal (en este caso, la cavidad abdominal), y la palabra mesentérico se refiere al sistema digestivo.

Figura 2. Conexiones simpáticas y ganglios en cadena. El axón de una neurona simpática central en la médula espinal puede proyectarse a la periferia de varias maneras. (a) La fibra puede proyectarse hacia el ganglio del mismo nivel y hacer sinapsis en una neurona ganglionar. (b) Una rama puede proyectarse a un ganglio más superior o inferior de la cadena. (c) Una rama puede proyectarse a través de la rama blanca comunicante, pero no terminar en una neurona ganglionar de la cadena. En su lugar, se proyecta a través de uno de los nervios esplácnicos a un ganglio colateral o a la médula suprarrenal (no aparece en la imagen).
Un axón de la neurona central que se proyecta a un ganglio simpático se denomina fibra o neurona preganglionar, y representa la salida del SNC hacia el ganglio. Dado que los ganglios simpáticos son adyacentes a la columna vertebral, las fibras simpáticas preganglionares son relativamente cortas y están mielinizadas. Una fibra postganglionar -el axón de una neurona ganglionar que se proyecta al efector diana- representa la salida de un ganglio que influye directamente en el órgano.
Comparadas con las fibras preganglionares, las fibras simpáticas postganglionares son largas debido a la distancia relativamente mayor desde el ganglio al efector diana. Estas fibras no están mielinizadas. (Obsérvese que el término «neurona postganglionar» puede utilizarse para describir la proyección desde un ganglio hasta la diana. El problema de este uso es que el cuerpo celular está en el ganglio y sólo la fibra es postganglionar. Normalmente, el término neurona se aplica a toda la célula.)
Un tipo de fibra simpática preganglionar no termina en un ganglio. Son los axones de las neuronas simpáticas centrales que se proyectan a la médula suprarrenal, la porción interior de la glándula suprarrenal. Estos axones todavía se denominan fibras preganglionares, pero el objetivo no es un ganglio. La médula suprarrenal libera moléculas de señalización en el torrente sanguíneo, en lugar de utilizar los axones para comunicarse con las estructuras objetivo. Las células de la médula suprarrenal con las que entran en contacto las fibras preganglionares se denominan células cromafines. Estas células son células neurosecretoras que se desarrollan a partir de la cresta neural junto con los ganglios simpáticos, lo que refuerza la idea de que la glándula es, funcionalmente, un ganglio simpático.
Las proyecciones de la división simpática del sistema nervioso autónomo divergen ampliamente, lo que da lugar a una amplia influencia del sistema en todo el cuerpo. Como respuesta a una amenaza, el sistema simpático aumentaría el ritmo cardíaco y la frecuencia respiratoria y haría que el flujo sanguíneo hacia el músculo esquelético aumentara y el flujo sanguíneo hacia el sistema digestivo disminuyera. La secreción de las glándulas sudoríparas también debería aumentar como parte de una respuesta integrada.
Se va a requerir que todos esos cambios fisiológicos ocurran juntos para huir de la leona cazadora, o el equivalente moderno. Esta divergencia se observa en los patrones de ramificación de las neuronas simpáticas preganglionares: una sola neurona simpática preganglionar puede tener entre 10 y 20 objetivos. Un axón que sale de una neurona central del cuerno lateral en la médula espinal toracolumbar pasará por la rama blanca comunicante y entrará en la cadena simpática, donde se ramificará hacia una variedad de objetivos. En el nivel de la médula espinal en el que la fibra simpática preganglionar sale de la médula espinal, una rama hará sinapsis en una neurona del ganglio de la cadena adyacente.
Algunas ramas se extenderán hacia arriba o hacia abajo a un nivel diferente de los ganglios de la cadena. Otras ramas atravesarán los ganglios de la cadena y se proyectarán a través de uno de los nervios esplácnicos hasta un ganglio colateral. Por último, algunas ramas pueden proyectarse a través de los nervios esplácnicos hasta la médula suprarrenal. Todas estas ramas significan que una neurona preganglionar puede influir en diferentes regiones del sistema simpático de forma muy amplia, actuando sobre órganos ampliamente distribuidos.
División parasimpática del sistema nervioso autónomo
La división parasimpática del sistema nervioso autónomo recibe su nombre porque sus neuronas centrales se encuentran a ambos lados de la región toracolumbar de la médula espinal (para- = «al lado» o «cerca»). El sistema parasimpático también puede denominarse sistema craneosacro (o de salida) porque las neuronas preganglionares se localizan en núcleos del tronco cerebral y en el cuerno lateral de la médula espinal sacra.
Las conexiones, o «circuitos», de la división parasimpática son similares a la disposición general de la división simpática con algunas diferencias específicas (Figura 3). Las fibras preganglionares de la región craneal viajan por los nervios craneales, mientras que las fibras preganglionares de la región sacra viajan por los nervios espinales. Los objetivos de estas fibras son los ganglios terminales, que se encuentran cerca -o incluso dentro- del efector objetivo. Estos ganglios se denominan a menudo ganglios intramurales cuando se encuentran dentro de las paredes del órgano diana. La fibra postganglionar se proyecta desde los ganglios terminales a una corta distancia del efector diana, o al tejido diana específico dentro del órgano. Si se comparan las longitudes relativas de los axones en el sistema parasimpático, las fibras preganglionares son largas y las postganglionares son cortas porque los ganglios están cerca -y a veces dentro- de los efectores diana.
El componente craneal del sistema parasimpático se basa en núcleos particulares del tronco cerebral. En el cerebro medio, el núcleo Eddinger-Westphal forma parte del complejo oculomotor, y los axones de esas neuronas viajan con las fibras del nervio oculomotor (nervio craneal III) que inervan los músculos extraoculares. Las fibras parasimpáticas preganglionares del III nervio craneal terminan en el ganglio ciliar, situado en la órbita posterior. Las fibras parasimpáticas postganglionares se proyectan al músculo liso del iris para controlar el tamaño de la pupila. En la médula superior, los núcleos salivales contienen neuronas con axones que se proyectan a través de los nervios faciales y glosofaríngeos a los ganglios que controlan las glándulas salivales. La producción de lágrimas está influenciada por las fibras parasimpáticas del nervio facial, que activan un ganglio y, en última instancia, la glándula lagrimal (lágrima).
Las neuronas del núcleo dorsal del nervio vago y del núcleo ambiguo se proyectan a través del nervio vago (nervio craneal X) a los ganglios terminales de las cavidades torácica y abdominal. Las fibras preganglionares parasimpáticas influyen principalmente en el corazón, los bronquios y el esófago de la cavidad torácica y en el estómago, el hígado, el páncreas, la vesícula biliar y el intestino delgado de la cavidad abdominal. Las fibras postganglionares de los ganglios activados por el nervio vago suelen incorporarse a la estructura del órgano, como el plexo mesentérico de los órganos del tubo digestivo y los ganglios intramurales.

Figura 3. Conexiones de la división parasimpática del sistema nervioso autónomo Las neuronas de los núcleos del tallo cerebral, o del cuerno lateral de la médula espinal sacra, se proyectan a los ganglios terminales cerca o dentro de los diversos órganos del cuerpo. Los axones de estas neuronas ganglionares se proyectan entonces a corta distancia hacia esos efectores objetivo.
Señalización química en el sistema nervioso autónomo
Cuando una neurona autonómica se conecta con un objetivo, se produce una sinapsis. La señal eléctrica del potencial de acción provoca la liberación de una molécula de señalización, que se unirá a las proteínas receptoras de la célula diana. Las sinapsis del sistema autónomo se clasifican como colinérgicas, lo que significa que se libera acetilcolina (ACh), o adrenérgicas, lo que significa que se libera norepinefrina. Los términos colinérgico y adrenérgico se refieren no sólo a la molécula de señalización que se libera, sino también a la clase de receptores a los que se une cada uno.
El sistema colinérgico incluye dos clases de receptores: el receptor nicotínico y el receptor muscarínico. Ambos tipos de receptores se unen a la ACh y provocan cambios en la célula diana. El receptor nicotínico es un canal de cationes cerrado por un ligando y el receptor muscarínico es un receptor acoplado a proteínas G. Los receptores reciben su nombre y se diferencian por otras moléculas que se unen a ellos. Mientras que la nicotina se une al receptor nicotínico, y la muscarina se une al receptor muscarínico, no hay reactividad cruzada entre los receptores. La situación es similar a la de las cerraduras y las llaves.
Imagina dos cerraduras -una para un aula y otra para una oficina- que se abren con dos llaves distintas. La llave del aula no abrirá la puerta de la oficina y la llave de la oficina no abrirá la puerta del aula. Esto es similar a la especificidad de la nicotina y la muscarina para sus receptores. Sin embargo, una llave maestra puede abrir múltiples cerraduras, como una llave maestra para el Departamento de Biología que abre tanto la puerta del aula como la del despacho. Esto es similar a la ACh que se une a ambos tipos de receptores. Las moléculas que definen estos receptores no son cruciales; son simplemente herramientas que los investigadores pueden utilizar en el laboratorio. Estas moléculas son exógenas, lo que significa que se fabrican fuera del cuerpo humano, por lo que un investigador puede utilizarlas sin que se produzcan resultados endógenos confusos (resultados causados por las moléculas producidas en el cuerpo).
El sistema adrenérgico también tiene dos tipos de receptores, denominados receptor alfa (α) adrenérgico y receptor beta (β) adrenérgico. A diferencia de los receptores colinérgicos, estos tipos de receptores no se clasifican por los fármacos que pueden unirse a ellos. Todos ellos son receptores acoplados a proteínas G. Hay tres tipos de receptores α-adrenérgicos, denominados α1, α2 y α3, y hay dos tipos de receptores β-adrenérgicos, denominados β1 y β2. Un aspecto adicional del sistema adrenérgico es que existe una segunda molécula de señalización denominada epinefrina. La diferencia química entre la norepinefrina y la epinefrina es la inclusión de un grupo metilo (CH3) en la epinefrina. El prefijo «nor-» se refiere en realidad a esta diferencia química, en la que falta un grupo metilo.
El término adrenérgico debería recordarle la palabra adrenalina, que se asocia con la respuesta de lucha o huida descrita al principio del capítulo. Adrenalina y epinefrina son dos nombres para la misma molécula. La glándula suprarrenal (en latín, ad- = «encima de»; renal = «riñón») segrega adrenalina. La terminación «-ine» se refiere a que la sustancia química se deriva, o se extrae, de la glándula suprarrenal. Una construcción similar del griego en lugar del latín da lugar a la palabra epinefrina (epi- = «encima de»; nephr- = «riñón»). En el uso científico, se prefiere epinefrina en Estados Unidos, mientras que adrenalina se prefiere en Gran Bretaña, porque «adrenalina» fue en su día un nombre de medicamento registrado y patentado en Estados Unidos. Aunque el fármaco ya no se vende, la convención de referirse a esta molécula con los dos nombres diferentes persiste. Del mismo modo, la norepinefrina y la noradrenalina son dos nombres para la misma molécula.
Habiendo entendido los sistemas colinérgico y adrenérgico, su papel en el sistema autónomo es relativamente sencillo de comprender. Todas las fibras preganglionares, tanto simpáticas como parasimpáticas, liberan ACh. Todas las neuronas ganglionares -los objetivos de estas fibras preganglionares- tienen receptores nicotínicos en sus membranas celulares. El receptor nicotínico es un canal de cationes cerrado por un ligando que provoca la despolarización de la membrana postsináptica. Las fibras parasimpáticas postganglionares también liberan ACh, pero los receptores en sus dianas son receptores muscarínicos, que son receptores acoplados a proteínas G y no causan exclusivamente la despolarización de la membrana postsináptica. Las fibras simpáticas postganglionares liberan norepinefrina, excepto las fibras que se proyectan a las glándulas sudoríparas y a los vasos sanguíneos asociados a los músculos esqueléticos, que liberan ACh (Tabla 1).
| Tabla 1 | ||
|---|---|---|
| Simpático | Parasimpático | |
| Preganglionar | Acetilcolina >Receptor nicotínico | Acetilcolina > receptor nicotínico | Postgangliónico | Norepinefrina > a o B-receptores adrenérgicos Acetilcolina >receptor muscarínico (asociado a las glándulas sudoríparas y a los vasos sanguíneos asociados a los músculos esqueléticos únicamente |
Acetilcolina > receptor muscarínico |
Las moléculas de señalización pueden pertenecer a dos grandes grupos. Los neurotransmisores se liberan en las sinapsis, mientras que las hormonas se liberan en el torrente sanguíneo. Estas son definiciones simplistas, pero pueden ayudar a aclarar este punto. La acetilcolina puede considerarse un neurotransmisor porque es liberada por los axones en las sinapsis. Sin embargo, el sistema adrenérgico presenta un desafío. Las fibras simpáticas postganglionares liberan norepinefrina, que puede considerarse un neurotransmisor. Pero la médula suprarrenal libera epinefrina y norepinefrina en la circulación, por lo que deben considerarse hormonas.
Lo que aquí se denomina sinapsis puede no ajustarse a la definición más estricta de sinapsis. Algunas fuentes se referirán a la conexión entre una fibra postganglionar y un efector objetivo como uniones neuroefectoras; los neurotransmisores, como se definió anteriormente, se llamarían neuromoduladores. La estructura de las conexiones postganglionares no son el típico bulbo terminal sináptico que se encuentra en la unión neuromuscular, sino que son cadenas de hinchamientos a lo largo de la longitud de una fibra postganglionar llamadas varicosidades (Figura 4).
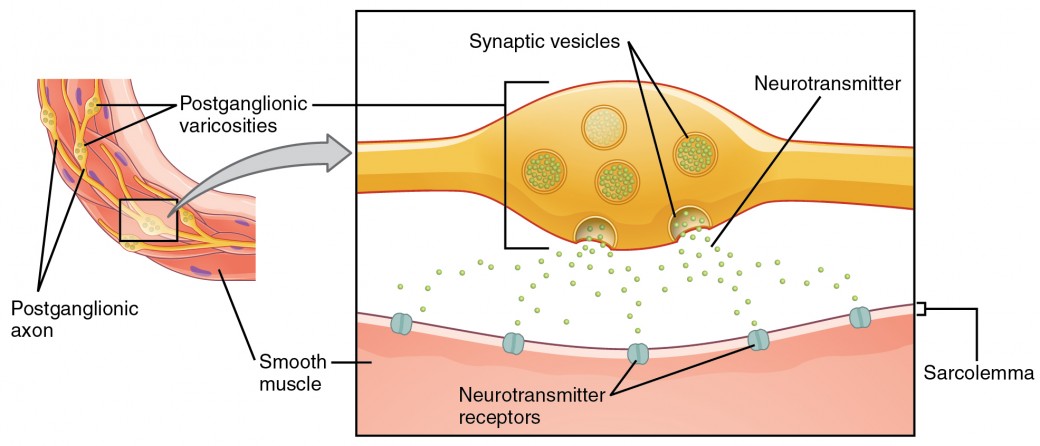
Figura 4. Varices autonómicas. La conexión entre las fibras autonómicas y los efectores diana no es la misma que la sinapsis típica, como la unión neuromuscular. En lugar de un bulbo terminal sináptico, se libera un neurotransmisor a partir de hinchazones a lo largo de la longitud de una fibra que hace una red extendida de conexiones en el efector objetivo.
El uso original del epíteto «lucha o huida» proviene de un científico llamado Walter Cannon que trabajó en Harvard en 1915. El concepto de homeostasis y el funcionamiento del sistema simpático habían sido introducidos en Francia en el siglo anterior. Cannon amplió la idea e introdujo la idea de que un animal responde a una amenaza preparándose para ponerse en pie y luchar o huir. La naturaleza de esta respuesta fue explicada a fondo en un libro sobre la fisiología del dolor, el hambre, el miedo y la ira.
Cuando los estudiantes aprenden sobre el sistema simpático y la respuesta de lucha o huida, a menudo se detienen y se preguntan sobre otras respuestas. Si usted se enfrentara a una leona que corre hacia usted, tal como se ilustra al principio de este capítulo, ¿correría o se mantendría firme? Algunas personas dirían que se quedarían paralizadas y no sabrían qué hacer. Entonces, ¿no hay realmente algo más en lo que hace el sistema autónomo que luchar, huir, descansar o digerir? ¿Qué pasa con el miedo y la parálisis ante una amenaza?
El epíteto común de «lucha o huida» se está ampliando para ser «lucha, huida o susto» o incluso «lucha, huida, susto o congelación». La aportación original de Cannon era una frase pegadiza para expresar parte de lo que hace el sistema nervioso en respuesta a una amenaza, pero es incompleta. El sistema simpático es responsable de las respuestas fisiológicas a los estados emocionales. Se puede decir que el nombre «simpático» significa eso (sym- = «junto»; -pathos = «dolor», «sufrimiento» o «emoción»).
Mira este vídeo para aprender más sobre el sistema nervioso.
Como se describe en este vídeo, el sistema nervioso tiene una forma de lidiar con las amenazas y el estrés que está separada del control consciente del sistema nervioso somático. El sistema proviene de una época en la que las amenazas tenían que ver con la supervivencia, pero en la era moderna, estas respuestas se convierten en parte del estrés y la ansiedad. Este vídeo describe cómo el sistema autónomo es sólo una parte de la respuesta a las amenazas, o a los factores de estrés. ¿Qué otro sistema de órganos interviene y qué parte del cerebro coordina los dos sistemas para la respuesta completa, incluyendo la epinefrina (adrenalina) y el cortisol?