Evaluación |Biopsicología |Comparativa |Cognitiva |Desarrollo |Lenguaje |Diferencias individuales. |Personalidad |Filosofía |Social |
Métodos |Estadística |Clínica |Educativa |Industrial |Profesional |Picología mundial |
Biológica:Genética del comportamiento – Psicología evolutiva – Neuroanatomía – Neuroquímica – Neuroendocrinología – Neurociencia – Psiconeuroinmunología – Psicología fisiológica – Psicofarmacología(Índice, Esquema)
| Rod cell | |
|---|---|
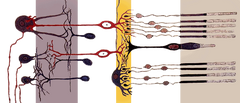
|
Localización | Retina |
| Función | Fotorreceptor de baja luminosidad |
| Morfología | Con forma de vara |
| Conexiones presinápticas | Ninguna | Conexiones postsinápticas | Células bipolares y células horizontales |
Las células rojas, o bastones, son células fotorreceptoras de la retina del ojo que pueden funcionar con una luz menos intensa que el otro tipo de fotorreceptor, los conos. Al ser más sensibles a la luz, los bastones son los responsables de la visión nocturna. Llamados así por su forma cilíndrica, los bastones se concentran en los bordes exteriores de la retina y se utilizan en la visión periférica. Hay unos 120 millones de células de bastón en la retina humana.
Una célula de bastón es lo suficientemente sensible como para responder a un solo fotón de luz, y es unas 100 veces más sensible a un solo fotón que los conos. Dado que los bastones necesitan menos luz para funcionar que los conos, son la principal fuente de información visual por la noche (visión escotópica). Los conos, en cambio, necesitan de decenas a cientos de fotones para activarse. Además, múltiples células de bastón convergen en una sola interneurona, recogiendo y amplificando las señales. Sin embargo, esta convergencia tiene un coste para la agudeza visual (o la resolución de la imagen), ya que la información agrupada de las múltiples células es menos nítida de lo que sería si el sistema visual recibiera la información de cada célula de bastón individualmente. La convergencia de las células de los bastones también tiende a hacer que la visión periférica sea muy sensible al movimiento, y es responsable del fenómeno de que un individuo vea algo vago por el rabillo del ojo.
Los bastones son un poco más estrechos que los conos pero tienen la misma base estructural. El pigmento se encuentra en el lado exterior, apoyado en el epitelio pigmentario. Este extremo contiene muchos discos apilados, probablemente por el plegado hacia dentro de la membrana limitante que rodea esta sección, lo que permite una mayor superficie para el pigmento visual y aumenta la eficacia de la absorción de la luz. Debido a que sólo tienen un tipo de pigmento sensible a la luz, en lugar de los tres tipos que tienen las células de los conos humanos, los bastones tienen poco o ningún papel en la visión del color.
Las células de los bastones también responden más lentamente a la luz que los conos, por lo que los estímulos que reciben se suman a lo largo de unos 100 milisegundos. Aunque esto hace que los bastones sean más sensibles a cantidades más pequeñas de luz, también significa que su capacidad para percibir cambios temporales, como imágenes que cambian rápidamente, es menos precisa que la de los conos.
Los experimentos realizados por George Wald y otros demostraron que los bastones son más sensibles a las longitudes de onda de la luz en torno a 498 nm (verde-azul), y son completamente insensibles a las longitudes de onda superiores a unos 640 nm (rojo). Este hecho es el responsable del efecto Purkinje, en el que los colores azules parecen más intensos en relación con los rojos en el crepúsculo, cuando los bastones toman el relevo como células responsables de la visión.
Al igual que los conos, las células de los bastones tienen un terminal sináptico, un segmento interno y un segmento externo. El terminal sináptico forma una sinapsis con otra neurona, por ejemplo una célula bipolar. Los segmentos interno y externo están conectados por un cilio. El segmento interno contiene orgánulos y el núcleo de la célula, mientras que el segmento externo del bastón (abreviado ROS), que apunta hacia la parte posterior del ojo, contiene los materiales que absorben la luz.
Respuesta a la luz
Anatomía de una célula de bastón
La activación de una sola molécula de rodopsina, el pigmento fotosensible de los bastones, puede provocar una gran reacción en la célula porque la señal se amplifica. Una vez activada, la rodopsina puede activar cientos de moléculas de transducina, cada una de las cuales activa a su vez una molécula de fosfodiesterasa, que puede descomponer más de mil moléculas de GMPc por segundo. Por lo tanto, los bastones pueden tener una gran respuesta a una pequeña cantidad de luz.
Como el componente retiniano de la rodopsina se deriva de la vitamina A, una deficiencia de ésta provoca un déficit del pigmento que necesitan las células de los bastones. En consecuencia, un menor número de células de los bastones es capaz de responder suficientemente en condiciones de oscuridad, y como las células de los conos están mal adaptadas para ver en la oscuridad, puede producirse ceguera. Se trata de la ceguera nocturna.
Revertir al estado de reposo
Los bastones hacen uso de tres mecanismos inhibitorios (mecanismos de retroalimentación negativa) para permitir una rápida reversión al estado de reposo tras un destello de luz.
En primer lugar, existe una rodopsina quinasa (RK) que fosforilaría la cola citosólica de la rodopsina activada en las múltiples serinas, inhibiendo parcialmente la activación de la transducina. Además, una proteína inhibidora -la arrestina- se une entonces a las rodopsinas fosforiladas para inhibir aún más la actividad de la rodopsina.
Mientras que la arrestina apaga la rodopsina, una proteína RGS (que funciona como una proteína activadora de GTPasas (GAPs)) lleva a la transducina (proteína G) a un estado «off» aumentando la tasa de hidrólisis del GTP ligado a GDP.
Además, como los canales sensibles al GMPc permiten no sólo la entrada de iones de sodio, sino también de iones de calcio, con la disminución de la concentración de GMPc, los canales sensibles al GMPc se cierran y reducen la entrada normal de iones de calcio. La disminución de la concentración de iones de calcio estimula las proteínas sensibles a los iones de calcio, que entonces activarían la guanilil ciclasa para reponer el GMPc, restaurando rápidamente su concentración original. El restablecimiento abre los canales sensibles al cGMP y provoca una despolarización de la membrana plasmática.
Desensibilización
Cuando los bastones se exponen a una alta concentración de fotones durante un periodo prolongado, se desensibilizan (se adaptan) al entorno.
Como la rodopsina es fosforilada por la rodopsina quinasa (un miembro de las GPCR quinasas (GRKs)), se une con alta afinidad a la arrestina. La arrestina unida puede contribuir al proceso de desensibilización al menos de dos maneras. En primer lugar, impide la interacción entre la proteína G y el receptor activado. En segundo lugar, sirve como proteína adaptadora para ayudar al receptor a la maquinaria de endocitosis dependiente de clatrina (para inducir la endocitosis mediada por el receptor).
Tabla
Comparación de las células de los bastones y los conos, de Kandel et al. (2000).
| Bastones | Conos |
|---|---|
| utilizados para la visión nocturna | utilizados para la visión diurna |
| no muy sensible a la luz; sensible sólo a la luz directa | |
| la pérdida provoca ceguera nocturna | la pérdida provoca ceguera legal |
| agudeza visual baja | agudeza visual alta; mejor resolución espacial |
| no presente en la fóvea | concentrada en la fóvea |
| respuesta lenta, a la luz, a los estímulos añadidos en el tiempo | respuesta rápida a la luz, pueden percibir cambios más rápidos en los estímulos |
| tienen más pigmento que los conos, por lo que pueden detectar menos luz | tienen menos pigmento que los bastones, requieren más luz para detectar las imágenes |
| los conjuntos de discosdiscos encerrados no están unidos a la membrana celular | los discos están unidos a la membrana externa |
| 20 veces más bastones que conos en la retina | |
| un tipo de pigmento fotosensible | tres tipos de pigmento fotosensible en los humanos | conferir visión acromática | conferir visión del color |
Referencia
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4th ed., pp.507-513. McGraw-Hill, Nueva York.
- Fisiología humana y mecanismos de la enfermedad de Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Molecular Biology of The Cell, 5th ed., pp.919-921. Garland Science. Cite error: Invalid
<ref>tag; name «Alberts» defined multiple times with different content
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4th ed., pp.507-513. McGraw-Hill, Nueva York.
.