Leerdoelen
- Noem de componenten die de sympathische en parasympathische reacties van het autonome zenuwstelsel genereren
- Licht de verschillen in output verbindingen binnen de twee divisies van het autonome zenuwstelsel
- Beschrijf de signaalmoleculen en receptoreiwitten die betrokken zijn bij de communicatie binnen de twee divisies van het autonome zenuwstelsel
Het zenuwstelsel kan worden onderverdeeld in twee functionele delen: het somatische zenuwstelsel en het autonome zenuwstelsel. De belangrijkste verschillen tussen de twee systemen komen tot uiting in de reacties die elk van beide teweegbrengt. Het somatisch zenuwstelsel veroorzaakt de samentrekking van de skeletspieren. Het autonome zenuwstelsel controleert hart- en gladde spieren, alsmede klierweefsel. Het somatische zenuwstelsel wordt geassocieerd met vrijwillige reacties (hoewel vele kunnen plaatsvinden zonder bewust bewustzijn, zoals ademhaling), en het autonome zenuwstelsel wordt geassocieerd met onwillekeurige reacties, zoals die in verband met homeostase.
Het autonome zenuwstelsel regelt veel van de inwendige organen door een evenwicht tussen twee aspecten, of divisies. In vergelijking met het endocriene systeem speelt het autonome zenuwstelsel een belangrijke rol bij de homeostatische mechanismen in het lichaam. De twee divisies van het autonome zenuwstelsel zijn de sympatische divisie en de parasympatische divisie. Het sympatische systeem wordt geassocieerd met de vecht-of-vluchtreactie, en de parasympathische activiteit wordt aangeduid met het epitheton rust en spijsvertering. Homeostase is het evenwicht tussen de twee systemen. Bij elke doel-effector bepaalt de dubbele innervatie de activiteit. Het hart bijvoorbeeld ontvangt verbindingen van zowel de sympatische als de parasympatische divisie. De ene zorgt ervoor dat de hartslag toeneemt, terwijl de andere juist zorgt dat de hartslag afneemt.
Bekijk deze video om meer te weten te komen over adrenaline en de vecht-of-vluchtreactie.
Wanneer iemand een adrenalinestoot zou hebben, komt meestal het beeld van bungeejumpers of parachutisten in je op. Maar adrenaline, ook bekend als epinefrine, is een belangrijke chemische stof bij het coördineren van de vecht-of-vluchtreactie van het lichaam. In deze video ziet u de fysiologie van de vecht-of-vluchtreactie, zoals een brandweerman die voor zich ziet. De reactie van zijn lichaam is het resultaat van de sympatische divisie van het autonome zenuwstelsel dat systeembrede veranderingen veroorzaakt terwijl het zich voorbereidt op extreme reacties. Welke twee veranderingen brengt adrenaline teweeg in de reactie van de skeletspieren?
Sympatische divisie van het autonome zenuwstelsel
Om te reageren op een bedreiging – vechten of vluchten – veroorzaakt het sympatische systeem uiteenlopende effecten doordat veel verschillende effectororganen samen worden geactiveerd voor een gemeenschappelijk doel. Er moet meer zuurstof worden ingeademd en aan de skeletspieren worden geleverd. Het ademhalingssysteem, het cardiovasculaire systeem en het bewegingsapparaat worden allemaal samen geactiveerd. Het zweten voorkomt dat het lichaam oververhit raakt door de overtollige warmte die ontstaat door het samentrekken van de spieren. Het spijsverteringsstelsel schakelt zich uit, zodat het bloed geen voedingsstoffen opneemt terwijl het juist zuurstof aan de skeletspieren moet leveren. Om al deze reacties te coördineren, lopen de verbindingen in het sympathische systeem uiteen van een beperkt gebied van het centrale zenuwstelsel (CZS) naar een breed scala van ganglia die naar de vele effectororganen tegelijk projecteren. De complexe reeks structuren die de output van het sympatische systeem samenstellen, maken het mogelijk dat deze ongelijksoortige effectoren samenkomen in een gecoördineerde, systemische verandering.
De sympatische afdeling van het autonome zenuwstelsel beïnvloedt de verschillende orgaansystemen van het lichaam via verbindingen die uit het thoracale en bovenste lumbale ruggenmerg komen. Het wordt het thoracolumbale systeem genoemd om deze anatomische basis weer te geven. Een centraal neuron in de laterale hoorn van een van deze spinale regio’s projecteert naar ganglia grenzend aan de wervelkolom via de ventrale spinale wortels.
De meeste ganglia van het sympatische systeem behoren tot een netwerk van sympatische keten ganglia dat langs de wervelkolom loopt. De ganglia zien eruit als een reeks clusters van neuronen die door axonale bruggen met elkaar verbonden zijn. Er zijn typisch 23 ganglia in de keten aan weerszijden van de wervelkolom. Drie komen overeen met het cervicale gebied, 12 met het thoracale gebied, vier met het lumbale gebied, en vier met het sacrale gebied. De cervicale en sacrale niveaus zijn niet rechtstreeks via de ruggenwortels met het ruggenmerg verbonden, maar via stijgende of dalende verbindingen via de bruggen binnen de keten.
Een diagram dat de verbindingen van het sympathische systeem weergeeft, lijkt enigszins op een schakelschema dat de elektrische verbindingen tussen verschillende recipiënten en apparaten weergeeft. In Figuur 1 zijn de “circuits” van het sympatische systeem opzettelijk vereenvoudigd.

Figuur 1. Verbindingen van de Sympatische Divisie van het Autonome Zenuwstelsel. Neuronen uit de laterale hoorn van het ruggenmerg (preganglionale neuronen) projecteren naar de kettingganglia aan weerszijden van de wervelkolom of naar collaterale (prevertebrale) ganglia die zich voor de wervelkolom in de buikholte bevinden. Axonen van deze ganglionnaire neuronen (postganglionaire vezels) projecteren zich vervolgens naar doeleffectoren in het gehele lichaam.
Om de analogie van het schakelschema voort te zetten: er zijn drie verschillende soorten “knooppunten” die binnen het sympathische systeem actief zijn (figuur 2). Het eerste type is het meest direct: de sympathische zenuw projecteert naar het ketenganglion op hetzelfde niveau als de doel-effector (het orgaan, weefsel of klier dat geïnnerveerd moet worden).
Een voorbeeld van dit type is spinale zenuw T1 die synapt met het T1 ketenganglion om de trachea te innerveren. De vezels van deze tak worden witte rami communicantes (enkelvoud = ramus communicans) genoemd; zij zijn gemyeliniseerd en worden daarom wit genoemd (zie figuur 2a). Het axon van het centrale neuron (de preganglionvezels weergegeven als een ononderbroken lijn) synapteert met het ganglionneuron (met de postganglionvezels weergegeven als een stippellijn). Dit neuron projecteert zich vervolgens naar een doel-effector – in dit geval de luchtpijp – via grijze rami communicantes, die ongemyeliniseerde axonen zijn.
In sommige gevallen bevinden de doel-effectoren zich superieur of inferieur aan het spinale segment waar de preganglionaire vezel ontspringt. Wat de “bedrading” betreft, vindt de synaps met het ganglionale neuron plaats in de ketenganglia superieur of inferieur aan de plaats van het centrale neuron. Een voorbeeld hiervan is de spinale zenuw T1 die het oog bedient. De spinale zenuw loopt door de keten omhoog tot hij het superieure cervicale ganglion bereikt, waar hij synapt met het postganglionale neuron (zie figuur 2b). De cervicale ganglia worden paravertebrale ganglia genoemd, gezien hun ligging naast de prevertebrale ganglia in de sympathische keten.
Niet alle axonen van de centrale neuronen eindigen in de ketenganglia. itionele takken van de ventrale zenuwwortel gaan verder door de keten en naar een van de collaterale ganglia als de n. splanchnicus major of n. splanchnicus lesser. Bijvoorbeeld, de grote splanchnische zenuw ter hoogte van T5 synapseert met een collaterale ganglia buiten de keten alvorens de verbinding te maken met de postganglionale zenuwen die de maag innerveren (zie Figuur 2c).
Collaterale ganglia, ook wel prevertebrale ganglia genoemd, bevinden zich anterieur aan de wervelkolom en ontvangen input van zowel splanchnische zenuwen als centrale sympathische neuronen. Zij worden in verband gebracht met de controle van organen in de buikholte, en worden ook beschouwd als deel van het enterische zenuwstelsel. De drie collaterale ganglia zijn het ganglion celiacus, het ganglion mesentericus superior en het ganglion mesentericus inferior (zie figuur 1). Het woord coeliakie is afgeleid van het Latijnse woord “coelom”, dat verwijst naar een lichaamsholte (in dit geval de buikholte), en het woord mesenterium verwijst naar het spijsverteringsstelsel.

Figuur 2. Sympatische verbindingen en ketenganglia. Het axon van een centraal sympathisch neuron in het ruggenmerg kan op een aantal verschillende manieren naar de periferie projecteren. (a) De vezel kan uitsteken naar het ganglion op hetzelfde niveau en synaps vertonen op een ganglionisch neuron. (b) Een tak kan projecteren naar een meer superieur of inferieur ganglion in de keten. (c) Een tak kan projecteren door de witte ramus communicans, maar niet eindigen op een ganglionisch neuron in de keten. In plaats daarvan projecteert hij via een van de splanchnische zenuwen naar een collateraal ganglion of het bijniermerg (niet afgebeeld).
Een axon van het centrale neuron dat naar een sympathisch ganglion projecteert, wordt een preganglionaire vezel of neuron genoemd, en vertegenwoordigt de output van het CZS naar het ganglion. Omdat de sympathische ganglia grenzen aan de wervelkolom, zijn de preganglionaire sympathische vezels relatief kort, en zijn ze gemyeliniseerd. Een postganglionaire vezel – het axon van een ganglionair neuron dat naar de doel-orgaan-orgaan projecteert – vertegenwoordigt de output van een ganglion dat rechtstreeks het orgaan beïnvloedt.
Vergeleken met de preganglionaire vezels zijn de postganglionaire sympathische vezels lang omdat de afstand van het ganglion tot de doel-orgaan-orgaan relatief groter is. Deze vezels zijn ongemyeliniseerd. (Merk op dat de term “postganglionisch neuron” kan worden gebruikt om de projectie van een ganglion naar het doelwit te beschrijven. Het probleem bij dat gebruik is dat het cellichaam zich in het ganglion bevindt, en dat alleen de vezel postganglionisch is. Typisch wordt de term neuron gebruikt voor de volledige cel.)
Eén type preganglionaire sympatische vezel eindigt niet in een ganglion. Dit zijn de axonen van centrale sympathische neuronen die projecteren naar de bijnier medulla, het inwendige gedeelte van de bijnier. Deze axonen worden nog steeds preganglionaire vezels genoemd, maar het doel is niet een ganglion. De bijniermerg geeft signaalmoleculen af in de bloedbaan, in plaats van axonen te gebruiken om te communiceren met doelstructuren. De cellen in het bijniermerg die door de preganglionaire vezels worden aangesproken, worden chromaffinecellen genoemd. Dit zijn neurosecretorische cellen die zich samen met de sympathische ganglia vanuit de neurale lijst ontwikkelen, wat het idee versterkt dat de klier functioneel een sympathisch ganglion is.
De projecties van de sympathische divisie van het autonome zenuwstelsel lopen sterk uiteen, wat resulteert in een brede invloed van het systeem in het hele lichaam. Als reactie op een dreiging zou het sympatische systeem de hartslag en de ademhaling verhogen en de bloedstroom naar de skeletspieren doen toenemen en de bloedstroom naar het spijsverteringsstelsel doen afnemen. De zweetklierafscheiding zou ook moeten toenemen als onderdeel van een geïntegreerde reactie.
Al deze fysiologische veranderingen moeten samen optreden om te kunnen vluchten voor de jagende leeuwin, of het moderne equivalent daarvan. Deze divergentie is te zien in de vertakkingspatronen van preganglionische sympathische neuronen – één preganglionisch sympathisch neuron kan 10-20 doelwitten hebben. Een axon dat een centraal neuron van de laterale hoorn in het thoracolumbale ruggenmerg verlaat, zal door de witte ramus communicans gaan en de sympatische keten binnengaan, waar het zich zal vertakken naar een verscheidenheid van doelwitten. Op het niveau van het ruggenmerg waar de preganglionaire sympatische vezel het ruggenmerg verlaat, zal een vertakking synapsen op een neuron in het aangrenzende ketting ganglion.
Sommige vertakkingen zullen zich naar boven of beneden uitstrekken naar een ander niveau van de ketting ganglia. Andere takken gaan door de ketenganglia en komen via een van de splanchnische zenuwen in een collateraal ganglion terecht. Tenslotte kunnen sommige takken via de splanchnische zenuwen naar de bijniermerg gaan. Al deze vertakkingen betekenen dat één preganglionisch neuron verschillende regio’s van het sympatische systeem zeer breed kan beïnvloeden, door in te werken op wijd verspreide organen.
Parasympathische afdeling van het autonome zenuwstelsel
De parasympathische afdeling van het autonome zenuwstelsel wordt zo genoemd omdat de centrale neuronen ervan zich aan weerszijden van het thoracolumbale gebied van het ruggenmerg bevinden (para- = “naast” of “dichtbij”). Het parasympathische systeem kan ook het craniosacrale systeem (of uitstroom) worden genoemd omdat de preganglionaire neuronen zich bevinden in kernen van de hersenstam en de laterale hoorn van het sacrale ruggenmerg.
De verbindingen, of “circuits”, van de parasympathische divisie zijn vergelijkbaar met de algemene lay-out van de sympatische divisie, met een paar specifieke verschillen (figuur 3). De preganglionaire vezels uit het craniale gebied lopen via craniale zenuwen, terwijl preganglionaire vezels uit het sacrale gebied via spinale zenuwen lopen. De doelwitten van deze vezels zijn terminale ganglia, die zich dichtbij of zelfs binnenin de doel-effector bevinden. Deze ganglia worden vaak intramurale ganglia genoemd wanneer zij zich binnen de wanden van het doelorgaan bevinden. De postganglionaire vezels projecteren zich vanuit de terminale ganglia over een korte afstand naar de doeleffector, of naar het specifieke doelweefsel binnen het orgaan. Bij vergelijking van de relatieve lengte van de axonen in het parasympathische systeem blijkt dat de preganglionaire vezels lang zijn en de postganglionaire vezels kort, omdat de ganglia zich dicht bij – en soms binnen – de doeleffectoren bevinden.
De craniale component van het parasympathische systeem is gebaseerd in bepaalde kernen van de hersenstam. In de middenhersenen maakt de kern Eddinger-Westphal deel uit van het oculomotorisch complex, en axonen van deze neuronen reizen samen met de vezels van de oculomotorische zenuw (nervus cranialis III) die de extraoculaire spieren inwendig bedienen. De preganglionaire parasympathische vezels van de n. cranialis III eindigen in het ganglion ciliaris, dat gelegen is in de achterste oogkas. De postganglionaire parasympatische vezels projecteren zich vervolgens naar de gladde spieren van de iris om de pupilgrootte te controleren. In de bovenste medulla bevatten de speekselkernen neuronen met axonen die via de gezichtszenuwen en de glossofaryngeale zenuwen naar de ganglia stromen die de speekselklieren controleren. De traanproductie wordt beïnvloed door parasympathische vezels in de nervus facialis, die een ganglion activeren, en uiteindelijk de lacrimale (traan)klier.
Neuronen in de dorsale nucleus van de nervus vagus en de nucleus ambiguus projecteren via de nervus vagus (hersenzenuw X) naar de terminale ganglia van de borstholte en buikholte. Parasympathische preganglionvezels beïnvloeden hoofdzakelijk het hart, de bronchiën en de slokdarm in de borstholte en de maag, lever, pancreas, galblaas en dunne darm in de buikholte. De postganglionaire vezels van de door de nervus vagus geactiveerde ganglia zijn vaak opgenomen in de structuur van het orgaan, zoals de plexus mesentericus van de organen van het spijsverteringskanaal en de intramurale ganglia.

Figuur 3. Neuronen vanuit kernen van de hersenstam of vanuit de laterale hoorn van het sacrale ruggenmerg, projecteren naar terminale ganglia nabij of in de verschillende organen van het lichaam. Axonen van deze ganglionale neuronen projecteren vervolgens de korte afstand naar deze doel-effectoren.
Chemische signalering in het autonome zenuwstelsel
Waar een autonoom neuron een verbinding maakt met een doelwit, is er sprake van een synaps. Het elektrische signaal van de actiepotentiaal veroorzaakt de afgifte van een signaalmolecuul, dat zich zal binden aan receptoreiwitten op de doelcel. Synapsen van het autonome systeem worden geclassificeerd als cholinerge synapsen, wat betekent dat er acetylcholine (ACh) vrijkomt, of adrenerge synapsen, wat betekent dat er noradrenaline vrijkomt. De termen cholinerge en adrenerge verwijzen niet alleen naar het signaalmolecuul dat vrijkomt, maar ook naar de klasse van receptoren die elk bindt.
Het cholinerge systeem omvat twee klassen receptoren: de nicotinereceptor en de muscarinereceptor. Beide receptortypes binden zich aan ACh en veroorzaken veranderingen in de doelcel. De nicotinereceptor is een ligand-gated kationkanaal en de muscarinereceptor is een G-eiwitgekoppelde receptor. De receptoren worden genoemd naar, en onderscheiden zich van elkaar door, andere moleculen die zich eraan binden. Terwijl nicotine aan de nicotinereceptor bindt en muscarine aan de muscarinereceptor, is er geen kruisreactiviteit tussen de receptoren. De situatie is vergelijkbaar met sloten en sleutels.
Stel je twee sloten voor – één voor een klaslokaal en één voor een kantoor – die met twee verschillende sleutels worden geopend. De sleutel van het klaslokaal zal de deur van het kantoor niet openen en de sleutel van het kantoor zal de deur van het klaslokaal niet openen. Dit is vergelijkbaar met de specificiteit van nicotine en muscarine voor hun receptoren. Een hoofdsleutel kan echter meerdere sloten openen, zoals een hoofdsleutel voor de afdeling Biologie die zowel de klaslokaal- als de kantoordeuren opent. Dit is vergelijkbaar met ACh dat zich bindt aan beide soorten receptoren. De moleculen die deze receptoren definiëren zijn niet van cruciaal belang – het zijn gewoon hulpmiddelen die onderzoekers in het laboratorium kunnen gebruiken. Deze moleculen zijn exogeen, wat betekent dat ze buiten het menselijk lichaam worden gemaakt, zodat een onderzoeker ze kan gebruiken zonder verwarrende endogene resultaten (resultaten die worden veroorzaakt door de moleculen die in het lichaam worden geproduceerd).
Het adrenerge systeem heeft ook twee soorten receptoren, de alfa (α)-adrenerge receptor en de bèta (β)-adrenerge receptor genoemd. In tegenstelling tot cholinerge receptoren zijn deze receptortypes niet ingedeeld volgens welke geneesmiddelen eraan kunnen binden. Het zijn allemaal G-eiwitgekoppelde receptoren. Er zijn drie typen α-adrenerge receptoren, die α1, α2 en α3 worden genoemd, en er zijn twee typen β-adrenerge receptoren, die β1 en β2 worden genoemd. Een bijkomend aspect van het adrenerge systeem is dat er een tweede signaalmolecule is die epinefrine wordt genoemd. Het chemische verschil tussen norepinefrine en epinefrine is de aanwezigheid van een methylgroep (CH3) in epinefrine. Het voorvoegsel “nor-” verwijst in feite naar dit chemische verschil, waarbij een methylgroep ontbreekt.
De term adrenerg moet je doen denken aan het woord adrenaline, dat wordt geassocieerd met de vecht-of-vluchtreactie die aan het begin van het hoofdstuk is beschreven. Adrenaline en epinefrine zijn twee namen voor dezelfde molecule. De bijnier (in het Latijn, ad- = “bovenop”; renaal = “nier”) scheidt adrenaline af. De uitgang “-ine” verwijst naar het feit dat de chemische stof wordt afgeleid, of geëxtraheerd, uit de bijnier. Een soortgelijke constructie uit het Grieks in plaats van het Latijn levert het woord epinefrine op (epi- = “boven”; nefr- = “nier”). In het wetenschappelijk gebruik wordt in de Verenigde Staten de voorkeur gegeven aan epinefrine, terwijl in Groot-Brittannië de voorkeur wordt gegeven aan adrenaline, omdat “adrenalin” ooit een geregistreerde, gepatenteerde geneesmiddelnaam was in de Verenigde Staten. Hoewel het geneesmiddel niet meer wordt verkocht, blijft de conventie bestaan om deze molecule met de twee verschillende namen aan te duiden. Evenzo zijn noradrenaline en norepinefrine twee namen voor dezelfde molecule.
Wanneer men het cholinerge en het adrenerge systeem heeft begrepen, is hun rol in het autonome systeem betrekkelijk eenvoudig te begrijpen. Alle preganglionaire vezels, zowel sympathische als parasympathische, geven ACh af. Alle ganglionnaire neuronen – de doelwitten van deze preganglionaire vezels – hebben nicotinereceptoren in hun celmembranen. De nicotinereceptor is een ligand-gated kationkanaal dat resulteert in depolarisatie van het postsynaptische membraan. De postganglionaire parasympathische vezels geven ook ACh af, maar de receptoren op hun doelwitten zijn muscarinereceptoren, die G-eiwitgekoppelde receptoren zijn en niet uitsluitend depolarisatie van het postsynaptische membraan veroorzaken. Postganglionische sympathische vezels geven noradrenaline af, met uitzondering van vezels die naar zweetklieren en naar bloedvaten in verband met skeletspieren lopen, die ACh afgeven (tabel 1).
| Tabel 1 | ||
|---|---|---|
| Sympatisch | Parasympatisch | |
| Preganglionisch | Acetylcholine > nicotinereceptor | Acetylcholine > nicotinereceptor |
| Postganglionische | Norepinefrine > a- of B-adrenerge receptoren Acetylcholine > muscarine receptor (alleen geassocieerd met zweetklieren en de bloedvaten geassocieerd met skeletspieren |
Acetylcholine > muscarine-receptor |
Signaalmoleculen kunnen tot twee grote groepen behoren. Neurotransmitters komen vrij bij synapsen, terwijl hormonen in de bloedbaan terechtkomen. Dit zijn simplistische definities, maar ze kunnen helpen om dit punt te verduidelijken. Acetylcholine kan als een neurotransmitter worden beschouwd omdat het door axonen bij synapsen wordt vrijgemaakt. Het adrenerge systeem vormt echter een uitdaging. Postganglionaire sympathische vezels geven noradrenaline af, dat als een neurotransmitter kan worden beschouwd. Maar het bijniermerg geeft epinefrine en noradrenaline af in de circulatie, zodat ze als hormonen moeten worden beschouwd.
Wat hier synapsen worden genoemd, voldoet misschien niet aan de meest strikte definitie van synapsen. Sommige bronnen verwijzen naar de verbinding tussen een postganglionaire vezel en een doel-effector als neuro-effector junctions; neurotransmitters, zoals hierboven gedefinieerd, zouden neuromodulatoren worden genoemd. De structuur van postganglionische verbindingen zijn niet de typische synaptische eindbol die wordt gevonden bij de neuromusculaire junctie, maar zijn eerder ketens van zwellingen langs de lengte van een postganglionische vezel die een varicositeit wordt genoemd (figuur 4).
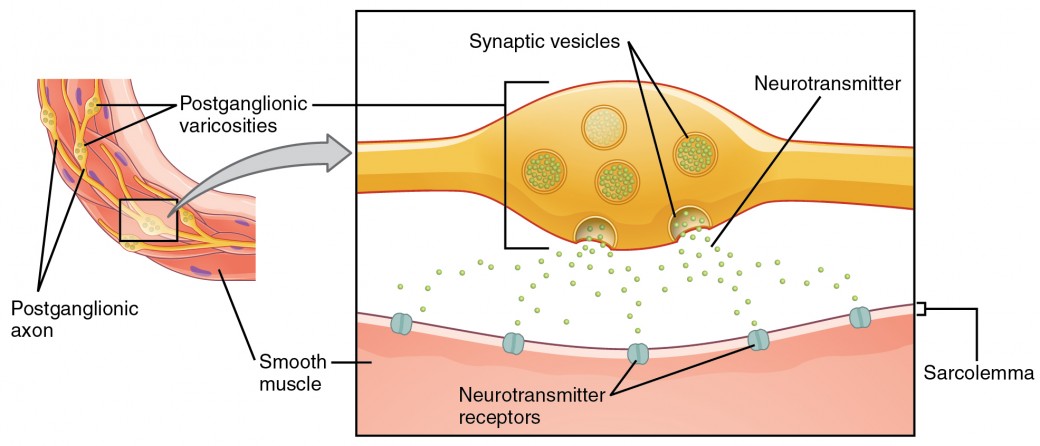
Figuur 4. Autonome varicositeiten. De verbinding tussen autonome vezels en doeleffectoren is niet dezelfde als de typische synaps, zoals de neuromusculaire junctie. In plaats van een synaptische eindbol, komt een neurotransmitter vrij uit zwellingen over de lengte van een vezel die een uitgebreid netwerk van verbindingen maakt in de doeleffector.
Elke Dag Connecties: Vechten of vluchten? Hoe zit het met angst en bevriezing?
Het oorspronkelijke gebruik van de uitdrukking “vecht of vlucht” is afkomstig van een wetenschapper genaamd Walter Cannon die in 1915 aan Harvard werkte. Het concept van homeostase en de werking van het sympathische systeem was in de vorige eeuw in Frankrijk geïntroduceerd. Cannon breidde het idee uit, en introduceerde het idee dat een dier op een bedreiging reageert door zich voor te bereiden om te blijven staan en te vechten of weg te rennen. De aard van deze reactie werd grondig uitgelegd in een boek over de fysiologie van pijn, honger, angst en woede.
Wanneer studenten leren over het sympatische systeem en de vecht-of-vlucht-reactie, staan ze vaak stil en vragen zich af hoe het zit met andere reacties. Als je geconfronteerd zou worden met een leeuwin die op je af komt rennen, zoals aan het begin van dit hoofdstuk is afgebeeld, zou je dan vluchten of zou je standhouden? Sommige mensen zouden zeggen dat ze zouden verstijven en niet weten wat te doen. Is het autonome systeem dan niet meer dan vechten, vluchten, rusten of verteren? Hoe zit het met angst en verlamming bij dreiging?
De gangbare term “vechten of vluchten” wordt uitgebreid tot “vechten, vluchten of bang zijn” of zelfs “vechten, vluchten, bang zijn of bevriezen”. De oorspronkelijke bijdrage van Cannon was een pakkende uitdrukking om iets uit te drukken van wat het zenuwstelsel doet als reactie op een bedreiging, maar hij is onvolledig. Het sympathisch systeem is verantwoordelijk voor de fysiologische reacties op emotionele toestanden. De naam “sympatisch” kan dat betekenen (sym- = “samen”; -pathos = “pijn,” “lijden,” of “emotie”).
Bekijk deze video om meer te weten te komen over het zenuwstelsel.
Zoals in deze video wordt beschreven, heeft het zenuwstelsel een manier om met bedreigingen en stress om te gaan die los staat van de bewuste controle van het somatische zenuwstelsel. Het systeem stamt uit een tijd waarin bedreigingen te maken hadden met overleven, maar in de moderne tijd zijn deze reacties onderdeel geworden van stress en angst. Deze video beschrijft hoe het autonome systeem slechts een deel is van de reactie op bedreigingen, of stressoren. Welk ander orgaansysteem is erbij betrokken, en welk deel van de hersenen coördineert de twee systemen voor de hele reactie, inclusief epinefrine (adrenaline) en cortisol?