Assessment | Biopsychology | Comparative |Cognitive | Developmental | Language | Individual differences |Persoonlijkheid |Filosofie |Sociaal |
Methodieken |Statistieken |Clinisch |Onderwijskundig |Industrieel |Professionele items |Wereldpsychologie |
Biologische:Gedragsgenetica – Evolutionaire psychologie – Neuroanatomie – Neurochemie – Neuroendocrinologie -Neurowetenschappen – Psychoneuroimmunologie – Fysiologische psychologie – Psychofarmacologie(Index, Outline)
| Rod cel | |
|---|---|
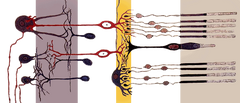
|
|
| Retina | |
| Functie | Laag licht fotoreceptor |
| Morfologie | staafvormig |
| Presynaptische verbindingen | Geen |
| Postsynaptische verbindingen | Bipolaire cellen en Horizontale cellen |
Roodcellen, of staafjes, zijn fotoreceptorcellen in het netvlies van het oog die kunnen functioneren bij minder intens licht dan het andere type fotoreceptor, de kegeltjescellen. Omdat ze lichtgevoeliger zijn, zijn staafjes verantwoordelijk voor het nachtzicht. De staafjes, zo genoemd naar hun cilindrische vorm, zijn geconcentreerd aan de buitenranden van het netvlies en worden gebruikt voor perifeer zicht. Er zijn ongeveer 120 miljoen staafcellen in het menselijk netvlies.
Een staafcel is gevoelig genoeg om op een enkel foton licht te reageren, en is ongeveer 100 keer gevoeliger voor een enkel foton dan kegeltjes. Omdat staafjes minder licht nodig hebben om te functioneren dan kegeltjes, zijn zij ’s nachts de primaire bron van visuele informatie (scotopisch zicht). Kegeltjescellen daarentegen hebben tientallen tot honderden fotonen nodig om geactiveerd te worden. Bovendien convergeren meerdere staafcellen op één enkel interneuron, waardoor de signalen worden verzameld en versterkt. Deze convergentie gaat echter ten koste van de gezichtsscherpte (of beeldresolutie), omdat de samengevoegde informatie van meerdere cellen minder onderscheidend is dan het geval zou zijn als het visuele systeem informatie van elke staafcel afzonderlijk zou ontvangen. De convergentie van staafcellen maakt het perifere zicht ook zeer gevoelig voor beweging, en is verantwoordelijk voor het verschijnsel dat iemand iets vaags uit zijn ooghoek ziet gebeuren.
Rods zijn iets smaller dan kegeltjes, maar hebben dezelfde structurele basis. Het pigment bevindt zich aan de buitenzijde, liggend op het pigmentepitheel. Dit uiteinde bevat veel gestapelde schijven, waarschijnlijk door het naar binnen vouwen van het begrenzende membraan dat dit gedeelte omgeeft, waardoor een groter oppervlak voor visueel pigment ontstaat en de efficiëntie van de lichtabsorptie toeneemt. Omdat zij slechts één type lichtgevoelig pigment hebben, in plaats van de drie typen die de menselijke kegeltjescellen hebben, spelen staafjes weinig of geen rol bij het kleurenzien.
Rode cellen reageren ook langzamer op licht dan kegeltjes, zodat de prikkels die zij ontvangen in ongeveer 100 milliseconden worden opgeteld. Dit maakt staafjes gevoeliger voor kleinere hoeveelheden licht, maar het betekent ook dat hun vermogen om veranderingen in de tijd waar te nemen, zoals snel veranderende beelden, minder nauwkeurig is dan dat van kegeltjes.
Uit experimenten van George Wald en anderen is gebleken dat staafjes het gevoeligst zijn voor golflengten van licht rond 498 nm (groen-blauw), en volledig ongevoelig voor golflengten langer dan ongeveer 640 nm (rood). Dit feit is verantwoordelijk voor het Purkinje-effect, waarbij blauwe kleuren in de schemering intenser lijken dan rode, wanneer de staafjes de taak overnemen van de cellen die verantwoordelijk zijn voor het gezichtsvermogen.
Net als kegels hebben staafcellen een synaptische terminal, een binnenste segment en een buitenste segment. Het synaptische uiteinde vormt een synaps met een ander neuron, bijvoorbeeld een bipolaire cel. Het binnenste en het buitenste segment zijn verbonden door een cilium. Het binnenste segment bevat organellen en de celkern, terwijl het buitenste segment van de staaf (afgekort tot ROS), dat naar de achterkant van het oog is gericht, de lichtabsorberende materialen bevat.
Reactie op licht
Anatomie van een staafcel
Activering van één enkele molecule rhodopsine, het lichtgevoelige pigment in staafjes, kan tot een grote reactie in de cel leiden omdat het signaal wordt versterkt. Eenmaal geactiveerd kan rhodopsine honderden transducinemoleculen activeren, die elk op hun beurt een fosfodiësterasemolecuul activeren, dat meer dan duizend cGMP-moleculen per seconde kan afbreken. Zo kunnen staafjes een grote respons geven op een kleine hoeveelheid licht.
Aangezien de retinale component van rhodopsine wordt afgeleid van vitamine A, veroorzaakt een tekort aan vitamine A een tekort aan het pigment dat de staafcellen nodig hebben. Als gevolg daarvan kunnen minder staafcellen voldoende reageren in donkere omstandigheden, en aangezien de kegeltjescellen slecht zijn aangepast voor zicht in het donker, kan blindheid het gevolg zijn. Dit is nachtblindheid.
Terugkeer naar de rusttoestand
Rods maken gebruik van drie remmende mechanismen (negatieve terugkoppelingsmechanismen) om een snelle terugkeer naar de rusttoestand na een lichtflits mogelijk te maken.
Eerst bestaat er een rhodopsinekinase (RK) die de cytosolische staart van het geactiveerde rhodopsine op de meervoudige serines zou fosforyleren, waardoor de activering van transducine gedeeltelijk wordt geremd. Bovendien bindt een remmend eiwit – arrestine – zich vervolgens aan de gefosforyleerde rhodopsines om de activiteit van het rhodopsine verder te remmen.
Terwijl arrestine rhodopsine uitschakelt, drijft een RGS-eiwit (dat functioneert als GTPase-activiating proteins(GAPs)) transducine (G-eiwit) in een “uit”-toestand door de snelheid van hydrolyse van het gebonden GTP tot GDP te verhogen.
Ook omdat de cGMP-gevoelige kanalen niet alleen de instroom van natriumionen, maar ook van calciumionen toelaten, worden met de verlaging van de concentratie cGMP de cGMP-gevoelige kanalen gesloten en vermindert de normale instroom van calciumionen. De daling van de concentratie calciumionen stimuleert de calciumion-gevoelige eiwitten, die dan het guanylyl cyclase activeren om het cGMP aan te vullen, waardoor de oorspronkelijke concentratie snel wordt hersteld. Het herstel opent de cGMP-gevoelige kanalen en veroorzaakt een depolarisatie van het plasmamembraan.
Desensibilisatie
Wanneer de staafjes langdurig aan een hoge concentratie fotonen worden blootgesteld, raken ze gedesensibiliseerd (aangepast) aan de omgeving.
Als rhodopsine wordt gefosforyleerd door rhodopsinekinase (een lid van de GPCR-kinases (GRK’s)), bindt het zich met hoge affiniteit aan het arrestine. Het gebonden arrestine kan op tenminste twee manieren bijdragen aan het desensibilisatieproces. Ten eerste verhindert het de interactie tussen het G-eiwit en de geactiveerde receptor. Ten tweede dient het als een adaptoreiwit om de receptor te helpen bij de clathrine-afhankelijke endocytose-machinerie (om receptorgemedieerde endocytose te induceren).
Tabel
Vergelijking van staaf- en kegelcellen, uit Kandel et al. (2000).
| Rods | Cones |
|---|---|
| gebruikt voor nachtzicht | gebruikt voor dagzicht |
| zeer lichtgevoelig; gevoelig voor strooilicht | niet erg lichtgevoelig; alleen gevoelig voor direct licht |
| verlies veroorzaakt nachtblindheid | verlies veroorzaakt rechtsblindheid |
| lage gezichtsscherpte | hoge gezichtsscherpte; betere ruimtelijke resolutie |
| niet aanwezig in fovea | geconcentreerd in fovea |
| langzame reactie, op licht, stimuli toegevoegd in de tijd | snelle reactie op licht, kan snellere veranderingen in stimuli waarnemen |
| hebben meer pigment dan kegeltjes, kunnen dus minder licht waarnemen | hebben minder pigment dan staafjes, hebben meer licht nodig om beelden waar te nemen |
| stapels van membraan-omsloten schijven zitten niet vast aan celmembraan | schijven zitten vast aan buitenmembraan |
| 20 keer meer staafjes dan kegeltjes in het netvlies | |
| één type lichtgevoelig pigment | drie soorten lichtgevoelig pigment bij de mens |
| zien achromatisch | zien kleuren |
Verwijzing
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Beginselen van de neurale wetenschap, 4e ed., pp.507-513. McGraw-Hill, New York.
- Human Physiology and Mechanisms of Disease door Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Molecular Biology of The Cell, 5th ed., pp.919-921. Garland Science. Cite error: Ongeldige
<ref>tag; naam “Alberts” meerdere malen gedefinieerd met verschillende inhoud
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Beginselen van de neurale wetenschap, 4e ed., pp.507-513. McGraw-Hill, New York.