Síntese de ácidos gordos ocorre no citoplasma e no retículo endoplasmático da célula e é quimicamente semelhante ao processo de beta-oxidação, mas com um par de diferenças chave. A primeira delas ocorre na preparação de substratos para as reacções que fazem crescer o ácido gordo. O transporte de acetil-CoA a partir das mitocôndrias ocorre quando estas começam a acumular-se. Duas moléculas podem desempenhar papéis no seu transporte para o citoplasma – citrato e acetilcarnitina. A união de oxaloacetato com acetil-CoA na mitocôndria cria o citrato que se move através da membrana, seguido pela acção da lisase do citrato no citoplasma da célula para libertar acetil-CoA e oxaloacetato. Além disso, quando a acetil-CoA livre se acumula na mitocôndria, pode combinar-se com a carnitina e ser transportada para o citoplasma.
Início com duas acetil-CoA, uma é convertida em malonil-CoA por carboxilação catalisada pela enzima acetil-CoA carboxilase (ACC), a única enzima reguladora da síntese de ácidos gordos (Figura \PageIndex{1}}). Em seguida, ambas as moléculas têm as suas porções de CoA substituídas por uma proteína transportadora conhecida como ACP (proteína transportadora de acilo) para formar acetil-ACP e malonil-ACP. A união de um acil-ACP gordo (neste caso, acetil-ACP) com malonil-ACP divide o carboxil que foi adicionado e cria o intermediário no canto superior direito na figura à esquerda.

Figure \(\PageIndex{1}}): Síntese de Ácido Gordo
A partir deste ponto, as reacções químicas assemelham-se às da oxidação beta invertida. Em primeiro lugar, a cetona é reduzida a um hidroxilo utilizando NADPH. Em contraste com o intermediário hidroxilado da oxidação beta, o intermediário beta aqui está na configuração D. Em seguida, a água é removida dos carbonos 2 e 3 do intermediário hidroxil para produzir uma molécula de ligação transduzida. Por último, a ligação dupla é hidrogenada para produzir um intermediário saturado. O processo cíclica com a adição de outro malonil-ACP à cadeia de crescimento até se produzir finalmente um intermediário com 16 carbonos (palmitoyl-CoA). Neste ponto, a síntese citoplasmática cessa.
Enzimas de Síntese de Ácido Gordo
Acetil-CoA carboxilase, que catalisa a síntese de malonil-CoA, é a única enzima regulada na síntese de ácidos gordos. A sua regulação envolve tanto o controlo alérgico como a modificação covalente. A enzima é conhecida por ser fosforilada tanto pela AMP Kinase como pela Proteína Kinase A. A desfosforilação é estimulada por fosfátases activadas pela ligação da insulina. A desfosforilação activa a enzima e favorece a sua montagem num polímero longo, enquanto a fosforilação inverte o processo. O citrato actua como um activador alostárico e pode também favorecer a polimerização. Palmitoyl-CoA aloéterically inactiva-o.
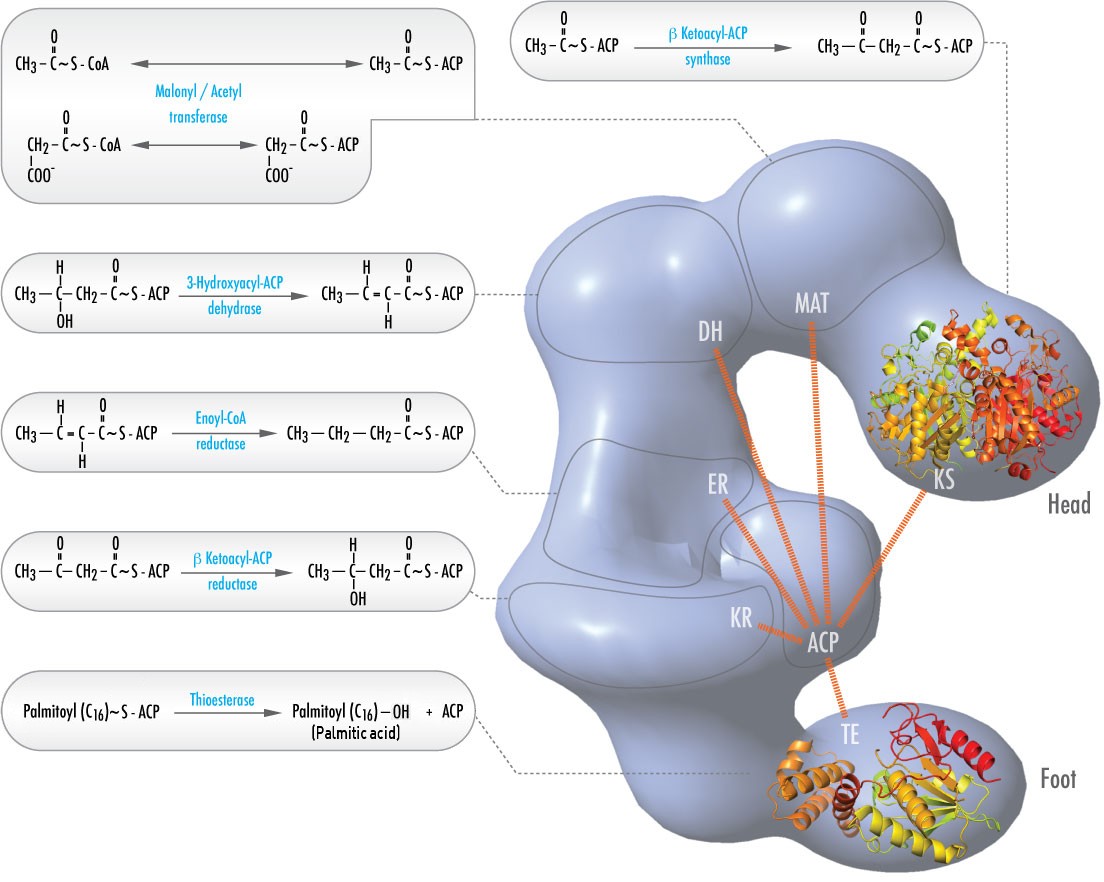
Em animais, seis diferentes actividades catalíticas necessárias para que as restantes acções catalíticas possam fazer palmitoyl-CoA estejam totalmente contidas num único complexo chamado Fatty Acid Synthase (Figura \PageIndex{2}). Estas incluem transacilases para troca de CoA com ACP em acetil-CoA e malonil-CoA; uma síntese para catalisar a adição das duas unidades de carbono das três unidades de malonil-ACP de carbono na primeira etapa do processo de alongamento; uma reductase para reduzir a cetona; uma desidrase para catalisar a remoção da água, e uma reductase para reduzir a dupla ligação trans. Em bactérias, estas actividades são encontradas em enzimas separadas e não fazem parte de um complexo.
Elongamento de Ácidos Gordos
Elongamento para fazer com que os ácidos gordos ultrapassem 16 carbonos ocorram no retículo endoplasmático e seja catalisado por enzimas descritas como elongases. As mitocôndrias também podem alongar os ácidos gordos, mas os seus materiais de partida são geralmente mais curtos do que 16 carbonos de comprimento. Os mecanismos em ambos os ambientes são semelhantes aos do citoplasma (um grupo malonil é utilizado para adicionar dois carbonos, por exemplo), mas o CoA está ligado aos intermediários, não ao ACP. Além disso, enquanto a síntese citoplasmática emprega o complexo de ácido gordo sintase (Figura \PageIndex{2}), as enzimas nestas organelas são separáveis e não fazem parte de um complexo.
Saturação de Ácidos Gordos
Ácidos gordos são sintetizados na forma saturada e a dessaturação ocorre mais tarde. Enzimas chamadas desaturases catalisam a formação de ligações duplas cis em ácidos gordos maduros. Estas enzimas são encontradas no retículo endoplasmático. Os animais são limitados nos ácidos gordos dessaturados que podem produzir, devido à incapacidade de catalisar reacções para além dos carbonos 9 e 10. Assim, os seres humanos podem fazer ácido oleico, mas não podem sintetizar ácido linoleico ou ácido linolénico. Consequentemente, estes dois devem ser fornecidos na dieta e são referidos como ácidos gordos essenciais.
Contribuintes
- p>r. Kevin Ahern e Dr. Indira Rajagopal (Universidade Estatal do Oregon)