Objectivos de Aprendizagem
- Nomear os componentes que geram as respostas simpáticas e parassimpáticas do sistema nervoso autónomo
- Explicar as diferenças de resultados ligações dentro das duas divisões do sistema nervoso autonómico
descrever as moléculas de sinalização e proteínas receptoras envolvidas na comunicação dentro das duas divisões do sistema nervoso autonómico
O sistema nervoso pode ser dividido em duas partes funcionais: o sistema nervoso somático e o sistema nervoso autonómico. As principais diferenças entre os dois sistemas são evidentes nas respostas que cada um produz. O sistema nervoso somático provoca a contracção dos músculos esqueléticos. O sistema nervoso autonómico controla os músculos cardíacos e lisos, bem como o tecido glandular. O sistema nervoso somático está associado a respostas voluntárias (embora muitas possam acontecer sem consciência, como a respiração), e o sistema nervoso autónomo está associado a respostas involuntárias, tais como as relacionadas com homeostasia.
O sistema nervoso autónomo regula muitos dos órgãos internos através de um equilíbrio de dois aspectos, ou divisões. Em relação ao sistema endócrino, o sistema nervoso autonómico é instrumental nos mecanismos homeostáticos do corpo. As duas divisões do sistema nervoso autonómico são a divisão simpática e a divisão parassimpática. O sistema simpático está associado à resposta de luta ou voo, e a actividade parassimpática é referida pelo epíteto de repouso e digestão. A homeostasia é o equilíbrio entre os dois sistemas. Em cada um dos efectores-alvo, a dupla inervação determina a actividade. Por exemplo, o coração recebe ligações tanto da divisão simpática como da parassimpática. Um faz aumentar a frequência cardíaca, enquanto o outro faz diminuir a frequência cardíaca.
Veja este vídeo para aprender mais sobre a adrenalina e a resposta de luta-ou-voo.
Quando se diz que alguém tem adrenalina, a imagem de bungee jumpers ou paraquedistas normalmente vem-me à cabeça. Mas a adrenalina, também conhecida como epinefrina, é um químico importante na coordenação da resposta do corpo à luta ou ao voo. Neste vídeo, observa-se o interior da fisiologia da resposta de combate ou de voo, tal como previsto para um bombeiro. A reacção do seu corpo é o resultado da divisão simpática do sistema nervoso autonómico, causando mudanças em todo o sistema à medida que este se prepara para respostas extremas. Que duas mudanças traz a adrenalina para ajudar a resposta do músculo esquelético?
Divisão Simpática do Sistema Nervoso Autónomo
Para responder a uma ameaça – lutar ou fugir – o sistema simpático causa efeitos divergentes, uma vez que muitos órgãos diferentes de efeito ou órgãos são activados em conjunto para um propósito comum. Mais oxigénio precisa de ser inalado e entregue ao músculo esquelético. Os sistemas respiratório, cardiovascular e músculo-esquelético são todos activados em conjunto. ialmente, o suor evita que o excesso de calor proveniente da contracção muscular provoque o sobreaquecimento do corpo. O sistema digestivo desliga-se para que o sangue não esteja a absorver nutrientes quando deveria estar a fornecer oxigénio aos músculos esqueléticos. Para coordenar todas estas respostas, as ligações no sistema simpático divergem de uma região limitada do sistema nervoso central (SNC) para uma vasta gama de gânglios que se projectam simultaneamente para os muitos órgãos efetores. O complexo conjunto de estruturas que compõem a saída do sistema simpático torna possível que estes diferentes efectores se juntem numa mudança coordenada e sistémica.
A divisão simpática do sistema nervoso autonómico influencia os vários sistemas de órgãos do corpo através de ligações que emergem da medula torácica e espinal lombar superior. É referido como o sistema toracolombar para reflectir esta base anatómica. Um neurónio central no corno lateral de qualquer destas regiões vertebrais projecta-se para os gânglios adjacentes à coluna vertebral através das raízes vertebrais ventrais.
A maioria dos gânglios do sistema simpático pertence a uma rede de gânglios de cadeia simpática que corre ao longo da coluna vertebral. Os gânglios aparecem como uma série de grupos de neurónios ligados por pontes axonais. Existem tipicamente 23 gânglios na cadeia de ambos os lados da coluna vertebral. Três correspondem à região cervical, 12 estão na região torácica, quatro na região lombar e quatro correspondem à região sacral. Os níveis cervical e sacral não estão ligados à medula espinal directamente através das raízes espinais, mas através de ligações ascendentes ou descendentes através das pontes dentro da cadeia.
Um diagrama que mostra as ligações do sistema simpático é um pouco como um diagrama de circuito que mostra as ligações eléctricas entre diferentes receptáculos e dispositivos. Na Figura 1, os “circuitos” do sistema simpático são intencionalmente simplificados.

Figure 1. Ligações da Divisão Simpática do Sistema Nervoso Autonómico. Os neurónios do corno lateral da medula espinal (neurónios pré-ganglionares) projectam-se para os gânglios em cadeia em ambos os lados da coluna vertebral ou para os gânglios colaterais (prevertebrais) que estão anteriores à coluna vertebral na cavidade abdominal. Os eixos destes neurónios ganglionares (fibras pós-ganglionares) projectam-se depois para os efectores-alvo em todo o corpo.
Para continuar com a analogia do diagrama do circuito, existem três tipos diferentes de “junções” que operam dentro do sistema simpático (Figura 2). O primeiro tipo é mais directo: o nervo simpático projecta-se para o gânglio em cadeia ao mesmo nível que o effector alvo (o órgão, tecido ou glândula a ser interiorizada).
Um exemplo deste tipo é o nervo espinhal T1 que sinapsula com o gânglio em cadeia T1 para interiorizar a traqueia. As fibras deste ramo são chamadas rami comunicantes brancos (singular = ramus communicans); são mielinizadas e por isso referidas como brancas (ver Figura 2a). O axônio do neurônio central (a fibra pré-ganglionar mostrada como uma linha sólida) sinapses com o neurônio ganglionar (com a fibra pós-ganglionar mostrada como uma linha tracejada). Este neurónio projecta-se então para um efeito alvo – neste caso, os comunicantes traquéia-via rami cinzento, que são axónios não mielinizados.
Em alguns casos, os efectores alvo estão localizados acima ou abaixo do segmento espinhal no qual emerge a fibra préganglionica. Com respeito à “cablagem” envolvida, a sinapse com o neurónio ganglionar ocorre em cadeias de gânglios superiores ou inferiores à localização do neurónio central. Um exemplo disto é o nervo espinhal T1 que inerva o olho. O nervo espinhal segue através da cadeia até atingir o gânglio cervical superior, onde sinapsa com o neurónio pós-ganglionar (ver Figura 2b). Os gânglios cervicais são referidos como gânglios paravertebrais, dada a sua localização adjacente aos gânglios de pré-vertebral na cadeia simpática.
Nem todos os axónios dos neurónios centrais terminam na cadeia de gânglios. ramos icionais da raiz do nervo ventral continuam através da cadeia e sobre um dos gânglios colaterais como o nervo esplâncnico maior ou o nervo esplâncnico menor. Por exemplo, o maior nervo esplâncnico ao nível das sinapses T5 com um gânglio colateral fora da cadeia antes de fazer a ligação aos nervos pós-ganglionares que se encontram no interior do estômago (ver Figura 2c).
Gânglios colaterais, também chamados gânglios pré-vertebrais, situam-se antes da coluna vertebral e recebem entradas dos nervos esplâncnicos, bem como dos neurónios centrais simpáticos. Estão associados aos órgãos de controlo da cavidade abdominal, e são também considerados como fazendo parte do sistema nervoso entérico. Os três gânglios colaterais são o gânglio celíaco, o gânglio mesentérico superior, e o gânglio mesentérico inferior (ver Figura 1). A palavra celíaco deriva da palavra latina “coelom”, que se refere a uma cavidade corporal (neste caso, a cavidade abdominal), e a palavra mesentérica refere-se ao sistema digestivo.

Figure 2. Ligações Simpáticas e Ganglia em Cadeia. O axónio de um neurónio simpático central na medula espinal pode projectar-se para a periferia de várias maneiras diferentes. (a) A fibra pode projectar-se para o gânglio ao mesmo nível e sinapse num neurónio ganglionar. (b) Um ramo pode projectar-se para mais gânglio superior ou inferior na cadeia. (c) Um ramo pode projectar-se através do ramus communicans branco, mas não terminar num neurónio ganglionar da cadeia. Em vez disso, projecta-se através de um dos nervos esplâncnicos para um gânglio colateral ou para a medula adrenal (não ilustrado).
Um axónio do neurónio central que se projecta para um gânglio simpático é referido como uma fibra ou neurónio pré-ganglionar, e representa a saída do CNS para o gânglio. Como os gânglios simpáticos são adjacentes à coluna vertebral, as fibras simpáticas pré-ganglionares são relativamente curtas, e são mielinizadas. Uma fibra pós-ganglionar – o axônio de um neurônio ganglionar que projeta para o efetor-alvo – representa a saída de um gânglio que influencia diretamente o órgão.
Comparado com as fibras pré-ganglionares, as fibras simpáticas pós-ganglionares são longas devido à distância relativamente maior entre o gânglio e o efetor-alvo. Estas fibras são não mielinizadas. (Note-se que o termo “neurónio pós-ganglionar” pode ser usado para descrever a projecção de um gânglio para o alvo. O problema com essa utilização é que o corpo celular está no gânglio, e apenas a fibra é pós-ganglionar. Tipicamente, o termo neurónio aplica-se a toda a célula.)
Um tipo de fibra simpática pré-ganglionar não termina num gânglio. Estes são os axónios dos neurónios simpáticos centrais que se projectam para a medula adrenal, a porção interior da glândula adrenal. Estes axónios ainda são referidos como fibras pré-ganglionares, mas o alvo não é um gânglio. A medula adrenal liberta moléculas sinalizadoras na corrente sanguínea, em vez de utilizar axónios para comunicar com as estruturas alvo. As células da medula adrenal que são contactadas pelas fibras pré-ganglionares são chamadas células cromatográficas. Estas células são células neurosecretoras que se desenvolvem a partir da crista neural juntamente com os gânglios simpáticos, reforçando a ideia de que a glândula é, funcionalmente, um gânglio simpático.
As projecções da divisão simpática do sistema nervoso autonómico divergem amplamente, resultando numa ampla influência do sistema em todo o corpo. Como resposta a uma ameaça, o sistema simpático aumentaria o ritmo cardíaco e respiratório e faria aumentar o fluxo sanguíneo para o músculo esquelético e diminuir o fluxo sanguíneo para o sistema digestivo. A secreção das glândulas sudoríparas deveria também aumentar como parte de uma resposta integrada.
Todas essas alterações fisiológicas terão de ocorrer em conjunto para fugir da leoa de caça, ou do equivalente moderno. Esta divergência é observada nos padrões de ramificação dos neurónios simpáticos préganglionares – um único neurónio simpático préganglionar pode ter 10-20 alvos. Um axónio que deixa um neurónio central do corno lateral na medula espinal toracolombar passará através do ramus communicans branco e entrará na cadeia simpática, onde se ramificará em direcção a uma variedade de alvos. Ao nível da medula espinal em que a fibra simpática pré-ganglionar sai da medula espinal, um ramo irá sinapsear num neurónio na cadeia ganglionar adjacente.
Alguns ramos irão estender-se para cima ou para baixo até um nível diferente da cadeia ganglionar. Outros ramos passarão através dos gânglios em cadeia e projectar-se-ão através de um dos nervos esplâncnicos até um gânglio colateral. Finalmente, alguns ramos poderão projectar-se através dos nervos esplâncnicos até à medula adrenal. Todos estes ramos significam que um neurónio pré-ganglionar pode influenciar diferentes regiões do sistema nervoso simpático de forma muito ampla, actuando em órgãos amplamente distribuídos.
Divisão Parassimpática do Sistema Nervoso Autónomo
A divisão parassimpática do sistema nervoso autónomo é nomeada porque os seus neurónios centrais estão localizados em ambos os lados da região toracolombar da medula espinal (para- = “ao lado” ou “perto”). O sistema parassimpático também pode ser referido como o sistema craniossacral (ou fluxo) porque os neurónios pré-ganglionares estão localizados em núcleos do tronco cerebral e do corno lateral da medula sacral.
As ligações, ou “circuitos”, da divisão parassimpática são semelhantes à disposição geral da divisão simpática com algumas diferenças específicas (Figura 3). As fibras préganglionares da região craniana viajam em nervos cranianos, enquanto as fibras préganglionares da região sacral viajam em nervos espinhais. Os alvos destas fibras são os gânglios terminais, que se localizam perto – ou mesmo dentro – do efector alvo. Estes gânglios são frequentemente referidos como gânglios intramurais quando são encontrados dentro das paredes do órgão alvo. As fibras pós-ganglionares projectam-se a partir dos gânglios terminais a uma curta distância até ao efector alvo, ou até ao tecido alvo específico dentro do órgão. Comparando os comprimentos relativos dos axónios no sistema parassimpático, as fibras pré-ganglionares são longas e as fibras pós-ganglionares são curtas porque os gânglios estão próximos – e por vezes dentro dos efectores-alvo.
O componente craniano do sistema parassimpático é baseado em núcleos específicos do tronco cerebral. No meio do cérebro, o núcleo Eddinger-Westphal faz parte do complexo oculomotor, e os axónios desses neurónios viajam com as fibras do nervo oculomotor (nervo craniano III) que inervam os músculos extra-oculares. As fibras parassimpáticas pré-ganglionares dentro do nervo craniano III terminam no gânglio ciliar, que está localizado na órbita posterior. As fibras parassimpáticas pós-ganglionares projectam-se então para o músculo liso da íris para controlar o tamanho pupilar. Na medula superior, os núcleos salivares contêm neurónios com axónios que se projectam através dos nervos faciais e glossofaríngeos para os gânglios que controlam as glândulas salivares. A produção de lágrimas é influenciada por fibras parassimpáticas no nervo facial, que activam um gânglio, e por fim a glândula lacrimal (lacrimal).
Neurões no núcleo dorsal do nervo vago e o núcleo ambíguo projectam-se através do nervo vago (nervo craniano X) até aos gânglios terminais das cavidades torácica e abdominal. As fibras parassimpáticas pré-ganglionares influenciam principalmente o coração, os brônquios e o esófago na cavidade torácica e o estômago, fígado, pâncreas, vesícula biliar, e intestino delgado da cavidade abdominal. As fibras pós-ganglionares dos gânglios activadas pelo nervo vago são frequentemente incorporadas na estrutura do órgão, tais como o plexo mesentérico dos órgãos do tracto digestivo e os gânglios intramurais.

p>Figure 3. As ligações da Divisão Parassimpática dos Neurónios do Sistema Nervoso Autónomo a partir dos núcleos cérebro-tronco, ou a partir do corno lateral da medula sacral, projectam-se para os gânglios terminais próximos ou dentro dos vários órgãos do corpo. Os axónios destes neurónios ganglionares projectam então a curta distância até aos efectores-alvo.
Sinalização Química no Sistema Nervoso Autónomo
Onde um neurónio autónomo se liga a um alvo, há uma sinapse. O sinal eléctrico do potencial de acção provoca a libertação de uma molécula de sinalização, que se ligará às proteínas receptoras na célula alvo. As sinapses do sistema autonómico são classificadas como colinérgicas, o que significa que a acetilcolina (ACh) é libertada, ou adrenérgicas, o que significa que a norepinefrina é libertada. Os termos colinérgico e adrenérgico referem-se não só à molécula de sinalização que é libertada mas também à classe de receptores que cada um liga.
O sistema colinérgico inclui duas classes de receptor: o receptor nicotínico e o receptor muscarínico. Ambos os tipos de receptores ligam-se ao ACh e provocam alterações na célula alvo. O receptor nicotínico é um canal catiónico ligeiro e o receptor muscarínico é um receptor acoplado à proteína G. Os receptores são nomeados e diferenciados por outras moléculas que se ligam a eles. Enquanto que a nicotina se liga ao receptor nicotínico, e a muscarina se liga ao receptor muscarínico, não há reactividade cruzada entre os receptores. A situação é semelhante às fechaduras e chaves.
p>Imagine duas fechaduras – uma para uma sala de aula e outra para um escritório – que são abertas por duas chaves separadas. A chave da sala de aula não abrirá a porta do escritório e a chave do escritório não abrirá a porta da sala de aula. Isto é semelhante à especificidade da nicotina e da muscarina para os seus receptores. No entanto, uma chave mestra pode abrir várias fechaduras, tais como uma chave mestra para o Departamento de Biologia que abre tanto a porta da sala de aula como a do escritório. Isto é semelhante ao ACh que se liga a ambos os tipos de receptores. As moléculas que definem estes receptores não são cruciais – são simplesmente ferramentas para os investigadores utilizarem no laboratório. Estas moléculas são exógenas, o que significa que são feitas fora do corpo humano, pelo que um investigador pode utilizá-las sem qualquer resultado endógeno confuso (resultados causados pelas moléculas produzidas no corpo).
O sistema adrenérgico também tem dois tipos de receptores, denominados receptor alfa (α)-adrenérgico e receptor beta (β)-adrenérgico. Ao contrário dos receptores colinérgicos, estes tipos de receptores não são classificados através dos quais as drogas se podem ligar a eles. Todos eles são receptores acoplados à proteína G. Existem três tipos de receptores α-adrenérgicos, denominados α1, α2, e α3, e existem dois tipos de receptores β-adrenérgicos, denominados β1 e β2. Um aspecto itional do sistema adrenérgico é que existe uma segunda molécula de sinalização chamada epinefrina. A diferença química entre norepinefrina e epinefrina é a ição de um grupo metilo (CH3) na epinefrina. O prefixo “nor-” refere-se efectivamente a esta diferença química, na qual falta um grupo metilo.
O termo adrenérgico deve lembrar a palavra adrenalina, que está associada à resposta luta-ou-voo descrita no início do capítulo. A adrenalina e epinefrina são dois nomes para a mesma molécula. A glândula adrenalina (em latim, ad- = “em cima de”; renal = “rim”) segrega a adrenalina. A terminação “-ina” refere-se ao produto químico derivado, ou extraído, da glândula adrenal. Uma construção semelhante do grego em vez do latim resulta na palavra epinefrina (epi- = “em cima”; nefr- = “rim”). No uso científico, a epinefrina é preferida nos Estados Unidos, enquanto que a adrenalina é preferida na Grã-Bretanha, porque “adrenalina” foi outrora um nome de droga registado e patenteado nos Estados Unidos. Embora a droga já não seja vendida, a convenção de referência a esta molécula pelos dois nomes diferentes persiste. Da mesma forma, norepinefrina e noradrenalina são dois nomes para a mesma molécula.
P>A partir do momento em que se compreende o sistema colinérgico e adrenérgico, o seu papel no sistema autonómico é relativamente simples de compreender. Todas as fibras pré-ganglionares, tanto simpáticas como parassimpáticas, libertam ACh. Todos os neurónios ganglionares – os alvos destas fibras pré-ganglionares – têm receptores nicotínicos nas suas membranas celulares. O receptor nicotínico é um canal catiónico ligeiro que resulta na despolarização da membrana pós-sináptica. As fibras parassimpáticas pós-ganglionares também libertam ACh, mas os receptores nos seus alvos são receptores muscarínicos, que são receptores acoplados à proteína G e não causam exclusivamente a despolarização da membrana pós-sináptica. As fibras simpáticas pós-ganglionares libertam norepinefrina, excepto as fibras que se projectam para as glândulas sudoríparas e para os vasos sanguíneos associados aos músculos esqueléticos, que libertam ACh (Tabela 1).
| Tabela 1 | ||
|---|---|---|
| Sympathetic | Parasympathetic | |
| Preganglionic | Acetylcholine > receptor nicotínico | Acetylcholine > receptor nicotínico |
| Postganglionic | Norepinefrina > a ou B-receptores adrenérgicos Acetylcholine > muscarinic receptor (associado às glândulas sudoríparas e aos vasos sanguíneos associados apenas aos músculos esqueléticos |
Acetylcholine > muscarinic receptor |
As moléculas de sinalização podem pertencer a dois grandes grupos. Os neurotransmissores são libertados nas sinapses, enquanto que as hormonas são libertadas na corrente sanguínea. Estas são definições simplistas, mas podem ajudar a clarificar este ponto. A acetilcolina pode ser considerada um neurotransmissor porque é libertada por axónios nas sinapses. O sistema adrenérgico, no entanto, apresenta um desafio. As fibras simpáticas pós-ganglionares libertam norepinefrina, que pode ser considerada um neurotransmissor. Mas a medula adrenal liberta epinefrina e norepinefrina em circulação, pelo que devem ser consideradas hormonas.
O que aqui são referidas como sinapses pode não se enquadrar na definição mais estrita de sinapse. Algumas fontes referir-se-ão à ligação entre uma fibra pós-ganglionar e um effector-alvo como junções neuroefectoras; os neurotransmissores, como definidos acima, seriam chamados neuromoduladores. A estrutura das ligações pós-ganglionares não é o típico bulbo sináptico final que se encontra na junção neuromuscular, mas sim cadeias de inchaços ao longo do comprimento de uma fibra pós-ganglionar chamada varicosidade (Figura 4).
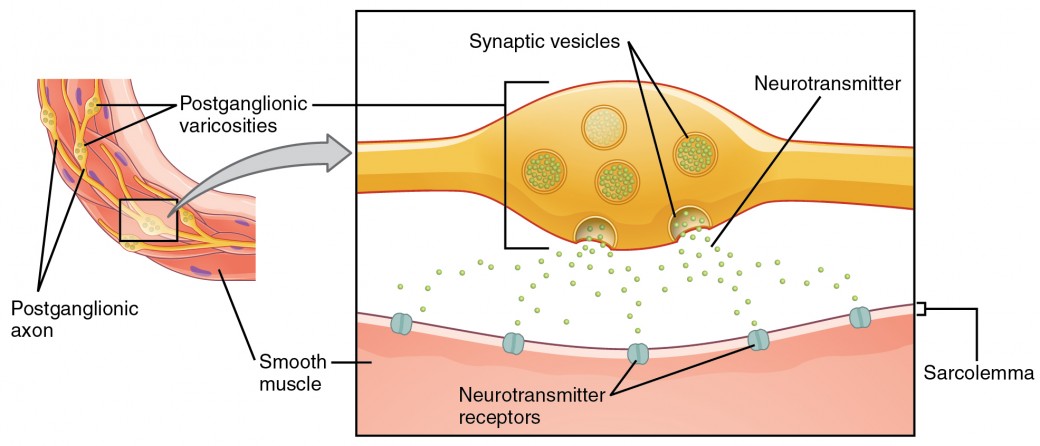
p>Figure 4. Varicosidades Autónomas. A ligação entre fibras autonómicas e efectores-alvo não é a mesma que a sinapse típica, tal como a junção neuromuscular. Em vez de um bulbo sináptico, um neurotransmissor é libertado de inchaços ao longo do comprimento de uma fibra que faz uma rede alargada de ligações no effector alvo.
Conexões diárias: Luta ou Voo? E o Fright and Freeze?
O uso original do epíteto “luta ou voo” vem de um cientista chamado Walter Cannon que trabalhou em Harvard em 1915. O conceito de homeostasia e o funcionamento do sistema simpático tinham sido introduzidos em França no século anterior. Cannon expandiu a ideia, e introduziu a ideia de que um animal responde a uma ameaça, preparando-se para ficar de pé e lutar ou fugir. A natureza desta resposta foi exaustivamente explicada num livro sobre a fisiologia da dor, fome, medo e raiva.
Quando os estudantes aprendem sobre o sistema simpático e a resposta de luta ou voo, muitas vezes param e questionam-se sobre outras respostas. Se fosse confrontado com uma leoa que corresse na sua direcção, tal como se encontra retratado no início deste capítulo, correria ou manteria a sua posição? Algumas pessoas diriam que ficariam congeladas e não saberiam o que fazer. Portanto, não há realmente mais do que lutar, voar, descansar, ou digerir no sistema autonómico. E o medo e a paralisia face a uma ameaça?
O epíteto comum de “lutar ou voar” está a ser alargado para ser “lutar, voar, ou assustar” ou mesmo “lutar, voar, assustar, ou congelar”. A contribuição original de Cannon era uma frase cativante para expressar um pouco do que o sistema nervoso faz em resposta a uma ameaça, mas está incompleta. O sistema simpático é responsável pelas respostas fisiológicas aos estados emocionais. O nome “simpático” pode ser dito para significar que (sym- = “juntos”; -pathos = “dor”, “sofrimento”, ou “emoção”).
Veja este vídeo para aprender mais sobre o sistema nervoso.
Como descrito neste vídeo, o sistema nervoso tem uma forma de lidar com ameaças e stress que está separada do controlo consciente do sistema nervoso somático. O sistema vem de uma época em que as ameaças eram sobre sobrevivência, mas na era moderna, estas respostas tornam-se parte do stress e da ansiedade. Este vídeo descreve como o sistema autônomo é apenas parte da resposta às ameaças, ou stressores. Que outro sistema de órgãos se envolve, e que parte do cérebro coordena os dois sistemas para toda a resposta, incluindo a epinefrina (adrenalina) e o cortisol?