Avaliação | Biopsicologia | Comparativo |Cognitivo |Devolutivo |Linguagem | Diferenças individuais |Filosofia |Filosofia |Social |br>Métodos |Estatísticas |Estatísticas |Clínicas |Educativas |Industriais |Profissionais |Psicologia Mundial |
Biológicas:Genética comportamental – Psicologia evolutiva – Neuroanatomia – Neuroquímica – Neuroendocrinologia – Neurociência – Psiconeuroimunologia – Psicologia Fisiológica – Psicofarmacologia(Índice), Esboço)
| Rod cell | |
|---|---|
|
|
| Localização | Retina |
| Função | Fotorreceptor de baixa luz |
| Morfologia | em forma de vara | Conexões pré-sinápticas | Nenhuma |
| Conexões pós-sinápticas | Células bipolares e células horizontais |
Células de Rod, ou hastes, são células fotorreceptoras na retina do olho que podem funcionar em luz menos intensa do que o outro tipo de células fotorreceptoras, células cone. Por serem mais sensíveis à luz, as varetas são responsáveis pela visão nocturna. Nomeadas pela sua forma cilíndrica, as hastes estão concentradas nos bordos exteriores da retina e são utilizadas na visão periférica. Existem cerca de 120 milhões de células de haste na retina humana.
Uma célula de haste é suficientemente sensível para responder a um único fotão de luz, e é cerca de 100 vezes mais sensível a um único fotão do que os cones. Como as hastes requerem menos luz para funcionar do que os cones, são portanto a principal fonte de informação visual durante a noite (visão escotópica). As células cónicas, por outro lado, necessitam de dezenas a centenas de fotões para serem activadas. Além disso, várias células de vara convergem num único interneurão, recolhendo e amplificando os sinais. No entanto, esta convergência tem um custo para a acuidade visual (ou resolução de imagem), porque a informação agrupada de múltiplas células é menos distinta do que seria se o sistema visual recebesse informação de cada célula de vara individualmente. A convergência de células de bastão também tende a tornar a visão periférica muito sensível ao movimento, e é responsável pelo fenómeno de um indivíduo ver algo vago ocorrer do canto do seu olho.
Rods são um pouco mais estreitos do que os cones, mas têm a mesma base estrutural. O pigmento está no lado exterior, deitado sobre o epitélio do pigmento. Esta extremidade contém muitos discos empilhados, provavelmente a partir da dobragem para dentro da membrana limitadora que envolve esta secção, permitindo uma área mais elevada para o pigmento visual e aumentando a eficiência da absorção da luz. Como têm apenas um tipo de pigmento sensível à luz, em vez dos três tipos que as células de cone humano têm, as hastes têm pouco, ou nenhum, papel na visão a cores.
Células de Rod também respondem mais lentamente à luz do que os cones, pelo que os estímulos que recebem são adicionados ao longo de cerca de 100 milissegundos. Embora isto torne as hastes mais sensíveis a pequenas quantidades de luz, significa também que a sua capacidade de detectar alterações temporais, tais como imagens de mudança rápida, é menos precisa do que a dos cones.
Experimentos de George Wald e outros mostraram que as hastes são mais sensíveis a comprimentos de onda de luz em torno de 498 nm (verde-azul), e são completamente insensíveis a comprimentos de onda superiores a cerca de 640 nm (vermelho). Este facto é responsável pelo efeito Purkinje, no qual as cores azuis aparecem mais intensas em relação aos vermelhos ao crepúsculo, quando as varas assumem o lugar das células responsáveis pela visão.
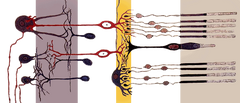
Células de bastão têm um terminal sináptico, um segmento interior, e um segmento exterior. O terminal sináptico forma uma sinapse com outro neurónio, por exemplo, uma célula bipolar. Os segmentos interno e externo são ligados por um cório. O segmento interno contém organelas e o núcleo da célula, enquanto o segmento externo da haste (abreviado para ROS), que é apontado para a parte de trás do olho, contém os materiais absorventes de luz.
Resposta à luz
Anatomia de uma Célula de Vara
p>A activação de uma única molécula de rodopsina, o pigmento fotossensível em varas, pode levar a uma grande reacção na célula porque o sinal é amplificado. Uma vez activada, a rodopsina pode activar centenas de moléculas de transducina, cada uma das quais por sua vez activa uma molécula de fosfodiesterase, que pode decompor mais de mil moléculas de cGMP por segundo. Assim, as varas podem ter uma grande resposta a uma pequena quantidade de luz.
Como o componente da retina da rodopsina é derivado da vitamina A, uma deficiência de vitamina A causa um défice no pigmento necessário às células da haste. Consequentemente, menos células de bastão são capazes de responder suficientemente em condições mais escuras, e como as células do cone estão mal adaptadas à visão no escuro, a cegueira pode resultar. Isto é cegueira nocturna.
Reverter para o estado de repouso
Rods fazem uso de três mecanismos inibidores (mecanismos de feedback negativo) para permitir um rápido retorno ao estado de repouso após um flash de luz.
P>Em primeiro lugar, existe uma rodopsina quinase (RK) que fosforilaria a cauda citosólica da rodopsina activada nas serinas múltiplas, inibindo parcialmente a activação da transducina. Além disso, uma proteína inibitória – detestin liga-se então às rhodopsinas fosforiladas para inibir ainda mais a actividade da rodopsina.
Enquanto o arrestin desliga a rodopsina, uma proteína RGS (funcionando como um GTP – proteínas activas (GAPs)) conduz a transducina (proteína G) para um estado “off”, aumentando a taxa de hidrólise do GTP limitado ao PIB.
Os canais sensíveis ao GTP também permitem não só o influxo de iões sódio, mas também de iões cálcio, com a diminuição da concentração de GTP, os canais sensíveis ao GTP são então fechados e reduzem o influxo normal de iões cálcio. A diminuição da concentração de iões de cálcio estimula as proteínas sensíveis aos iões de cálcio, que activariam então a guanylyl ciclase para reabastecer o GMPc, restaurando rapidamente a sua concentração original. A restauração abre os canais sensíveis ao cGMP e provoca uma despolarização da membrana plasmática.
Desensibilização
Quando as hastes são expostas a uma alta concentração de fotões durante um período prolongado, ficam dessensibilizadas (adaptadas) ao ambiente.
As rhodopsin é fosforizada por rhodopsin kinase (um membro da GPCR kinases(GRKs)), liga-se com grande afinidade à prisão. A prisão vinculada pode contribuir para o processo de dessensibilização de pelo menos duas formas. Primeiro, impede a interacção entre a proteína G e o receptor activado. Em segundo lugar, serve como uma proteína adaptadora para ajudar o receptor ao mecanismo de endocitose dependente da clatrina (para induzir a endocitose mediada pelo receptor).
Tabela
Comparação de células de barra e cone, de Kandel et al. (2000).
| Rods | Cones |
|---|---|
| usado para visão nocturna | usado para visão diurna |
| não muito sensível à luz; sensível apenas à luz directa | |
| perda causa cegueira legal | |
| baixa acuidade visual | alta acuidade visual; melhor resolução espacial |
| concentrada em fovea | |
| resposta rápida à luz, pode perceber mudanças mais rápidas nos estímulos | |
| tem menos pigmento do que as varas, necessitam de mais luz para detectar imagens | |
| stacks de membrana-Os discos fechados não estão ligados à membrana celular | discos estão ligados à membrana externa |
| um tipo de pigmento fotossensível | tr tipos de pigmentos fotossensíveis em humanos |
| confer visão acromática | confer visão colorida |
Referência
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4ª ed., pp.507-513. McGraw-Hill, Nova Iorque.
- Human Physiology and Mechanisms of Disease por Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Molecular Biology of The Cell, 5ª ed., pp.919-921. Garland Science. Citar erro: Inválido
<ref>tag; nome “Alberts” definido várias vezes com conteúdo diferente
- >li>Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4ª ed., pp.507-513. McGraw-Hill, New York.