Ocena | Biopsychologia | Porównawcza | Poznawcza | Rozwojowa | Językowa | Różnice indywidualne | Osobowość | Filozofia | Społeczne |
Metody | Statystyka |Kliniczne | Edukacyjne | Przemysłowe |Przedmioty zawodowe | Psychologia światowa |
Biologiczne:Genetyka behawioralna – Psychologia ewolucyjna – Neuroanatomia – Neurochemia – Neuroendokrynologia -Neuroscience – Psychoneuroimmunologia – Psychologia fizjologiczna – Psychofarmakologia(Indeks, Konspekt)
| Rod komórka | |
|---|---|
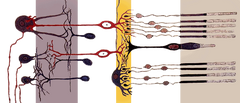
|
|
| Lokalizacja | Siatkówka |
| Funkcja | Fotoreceptor dla światła słabego |
| Morfologia | kształt pręcika |
| Połączenia presynaptyczne | Nie |
| Połączenia postsynaptyczne | Komórki dwubiegunowe i komórki horyzontalne |
Komórki pręcikowe, lub pręciki, są komórkami fotoreceptorowymi w siatkówce oka, które mogą funkcjonować w mniej intensywnym świetle niż drugi typ fotoreceptorów, komórki stożkowe. Ponieważ są one bardziej wrażliwe na światło, pręciki są odpowiedzialne za widzenie nocne. Nazwane tak ze względu na ich cylindryczny kształt, pręciki są skoncentrowane na zewnętrznych krawędziach siatkówki i są wykorzystywane w widzeniu peryferyjnym. W ludzkiej siatkówce znajduje się około 120 milionów komórek pręcikowych.
Komórka pręcikowa jest wystarczająco wrażliwa, aby reagować na pojedynczy foton światła, i jest około 100 razy bardziej wrażliwa na pojedynczy foton niż czopki. Ponieważ pręciki wymagają mniej światła do funkcjonowania niż czopki, są one głównym źródłem informacji wizualnej w nocy (widzenie skotopowe). Z drugiej strony, komórki stożkowe potrzebują od kilkudziesięciu do kilkuset fotonów, aby się uaktywnić. Dodatkowo, wiele komórek pręcikowych skupia się na jednym interneuronie, zbierając i wzmacniając sygnały. Zbieżność ta ma jednak negatywny wpływ na ostrość widzenia (rozdzielczość obrazu), ponieważ informacje pochodzące z wielu komórek są mniej wyraźne niż gdyby układ wzrokowy otrzymywał informacje z każdej komórki pręcikowej osobno. Zbieżność komórek pręcików powoduje, że widzenie peryferyjne jest bardzo wrażliwe na ruch i jest odpowiedzialne za zjawisko, w którym osoba widzi coś niewyraźnego w kąciku oka.
Pręciki są nieco węższe niż czopki, ale mają tę samą podstawę strukturalną. Barwnik znajduje się na zewnętrznej stronie, leżąc na nabłonku pigmentowym. Ten koniec zawiera wiele ułożonych dysków, prawdopodobnie z fałdowania do wewnątrz błony ograniczającej otaczającej tę część, co pozwala na większą powierzchnię dla pigmentu wzrokowego i zwiększa efektywność absorpcji światła. Ponieważ posiadają one tylko jeden rodzaj światłoczułego pigmentu, a nie trzy rodzaje, które posiadają ludzkie komórki czopkowe, pręciki odgrywają niewielką, jeśli w ogóle, rolę w widzeniu barwnym.
Komórki pręcikowe reagują również wolniej na światło niż czopki, więc bodźce, które otrzymują, sumują się w ciągu około 100 milisekund. To sprawia, że pręciki są bardziej wrażliwe na mniejsze ilości światła, ale oznacza również, że ich zdolność do wyczuwania zmian czasowych, takich jak szybko zmieniające się obrazy, jest mniej dokładna niż zdolność czopków.
Eksperymenty George’a Walda i innych wykazały, że pręciki są najbardziej wrażliwe na fale światła o długości około 498 nm (zielono-niebieskie) i są całkowicie niewrażliwe na fale dłuższe niż około 640 nm (czerwone). Fakt ten jest odpowiedzialny za efekt Purkinjego, w którym niebieskie kolory wydają się bardziej intensywne w stosunku do czerwonych o zmierzchu, kiedy to pręciki przejmują rolę komórek odpowiedzialnych za widzenie.
Jak czopki, komórki pręcikowe mają terminal synaptyczny, segment wewnętrzny i segment zewnętrzny. Terminal synaptyczny tworzy synapsę z innym neuronem, na przykład z komórką dwubiegunową. Segmenty wewnętrzny i zewnętrzny są połączone cewką. Segment wewnętrzny zawiera organelle i jądro komórkowe, podczas gdy segment zewnętrzny pręcika (w skrócie ROS), który jest skierowany w stronę tylnej części oka, zawiera materiały pochłaniające światło.
Reakcja na światło
Anatomy of a Rod Cell
Aktywacja pojedynczej cząsteczki rodopsyny, światłoczułego pigmentu w pręcikach, może prowadzić do dużej reakcji w komórce, ponieważ sygnał jest wzmacniany. Po aktywacji, rodopsyna może aktywować setki cząsteczek transducyny, z których każda z kolei aktywuje cząsteczkę fosfodiesterazy, która może rozłożyć ponad tysiąc cząsteczek cGMP na sekundę. W ten sposób pręciki mogą mieć dużą odpowiedź na małą ilość światła.
Jako że składnik siatkówkowy rodopsyny pochodzi z witaminy A, niedobór witaminy A powoduje deficyt pigmentu potrzebnego komórkom pręcików. W związku z tym mniej komórek pręcikowych jest w stanie odpowiednio reagować w ciemności, a ponieważ komórki czopkowe są słabo przystosowane do widzenia w ciemności, może dojść do ślepoty. Jest to ślepota nocna.
Powrót do stanu spoczynku
Różdżki wykorzystują trzy mechanizmy hamujące (mechanizmy ujemnego sprzężenia zwrotnego), aby umożliwić szybki powrót do stanu spoczynku po błysku światła.
Po pierwsze, istnieje kinaza rodopsyny (RK), która fosforyluje cytozolowy ogon aktywowanej rodopsyny na wielu serynach, częściowo hamując aktywację transducyny. Do ufosforylowanej rodopsyny przyłącza się również białko hamujące – aretenina, która dalej hamuje aktywność rodopsyny.
Podczas gdy aresztyna wyłącza rodopsynę, białko RGS (funkcjonujące jako białko aktywujące GTPazy (GAPs)) wprowadza transducynę (białko G) w stan „wyłączony” poprzez zwiększenie tempa hydrolizy związanego GTP do GDP.
Jak również kanały wrażliwe na cGMP umożliwiają nie tylko napływ jonów sodu, ale również jonów wapnia, wraz ze spadkiem stężenia cGMP kanały wrażliwe na cGMP są zamykane i zmniejszają normalny napływ jonów wapnia. Spadek stężenia jonów wapnia pobudza białka wrażliwe na jony wapnia, które następnie aktywują cyklazę guanylową do uzupełnienia cGMP, szybko przywracając jego pierwotne stężenie. Przywrócenie otwiera kanały wrażliwe na cGMP i powoduje depolaryzację błony plazmatycznej.
Odczulanie
Gdy pręciki są narażone na wysokie stężenie fotonów przez dłuższy czas, stają się odczulone (przystosowane) do środowiska.
Jako że rodopsyna jest fosforylowana przez kinazę rodopsyny (członek kinaz GPCR (GRKs)), wiąże się z wysokim powinowactwem do areteny. Związana arestyna może przyczyniać się do procesu desensytyzacji na co najmniej dwa sposoby. Po pierwsze, zapobiega ona interakcji pomiędzy białkiem G a aktywowanym receptorem. Po drugie, służy jako białko adaptorowe, które pomaga receptorowi dotrzeć do mechanizmów endocytozy zależnej od klatyny (w celu wywołania endocytozy pośredniczącej w receptorze).
Tabela
Porównanie komórek pręcikowych i czopkowych, z Kandel et al. (2000).
| Rod | Cones |
|---|---|
| używane do widzenia nocnego | używane do widzenia dziennego |
| bardzo wrażliwe na światło; wrażliwy na światło rozproszone | niezbyt wrażliwy na światło; wrażliwy tylko na światło bezpośrednie |
| utraty powodują ślepotę nocną | utraty powodują ślepotę prawną |
| niska ostrość widzenia | wysoka ostrość widzenia; lepsza rozdzielczość przestrzenna |
| nie występuje w fovea | koncentruje się w fovea |
| wolna reakcja, na światło, bodźce dodawane w czasie | szybka reakcja na światło, mogą odbierać szybsze zmiany w bodźcach |
| mają więcej pigmentu niż czopki, więc mogą wykryć mniej światła | mają mniej pigmentu niż pręciki, wymagają więcej światła do wykrywania obrazów |
| stosy błoniastych tarcztarczki są nieprzymocowane do błony komórkowej | tarczki są przymocowane do błony zewnętrznej |
| 20 razy więcej pręcików niż czopków w siatkówce | |
| jeden typ światłoczułego pigmentu | trzy typów pigmentu światłoczułego u ludzi |
| pozwalają na widzenie achromatyczne | pozwalają na widzenie barwne |
Referencje
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4th ed., pp.507-513. McGraw-Hill, New York.
- Human Physiology and Mechanisms of Disease by Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Molecular Biology of The Cell, 5th ed., pp.919-921. Garland Science. Cite error: Invalid
<ref>tag; name „Alberts” defined multiple times with different content
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4th ed., pp.507-513. McGraw-Hill, New York.
.