La sintesi degli acidi grassi avviene nel citoplasma e nel reticolo endoplasmatico della cellula ed è chimicamente simile al processo di beta-ossidazione, ma con un paio di differenze chiave. La prima di queste si verifica nella preparazione dei substrati per le reazioni che fanno crescere l’acido grasso. Il trasporto di acetil-CoA dai mitocondri avviene quando comincia ad accumularsi. Due molecole possono giocare un ruolo nello spostamento verso il citoplasma: il citrato e l’acetilcarnitina. L’unione dell’ossalacetato con l’acetil-CoA nel mitocondrio crea il citrato che si muove attraverso la membrana, seguito dall’azione della citrato liasi nel citoplasma della cellula per rilasciare acetil-CoA e ossalacetato. Inoltre, quando l’acetil-CoA libero si accumula nel mitocondrio, può combinarsi con la carnitina ed essere trasportato fuori nel citoplasma.
A partire da due acetil-CoA, uno è convertito in malonil-CoA dalla carbossilazione catalizzata dall’enzima acetil-CoA carbossilasi (ACC), l’unico enzima regolatore della sintesi degli acidi grassi (Figura \(\PageIndex{1}). Successivamente, entrambe le molecole hanno le loro porzioni di CoA sostituite da una proteina trasportatrice conosciuta come ACP (acyl-carrier protein) per formare acetil-ACP e malonil-ACP. L’unione di un acile grasso-ACP (in questo caso, acetil-ACP) con il malonil-ACP scinde il carbossile che è stato aggiunto e crea l’intermedio in alto a destra nella figura a sinistra.

Figura \(\PageIndex{1}): Sintesi degli acidi grassi
Da questo punto in avanti, le reazioni chimiche assomigliano a quelle della beta ossidazione invertita. In primo luogo, il chetone è ridotto a un idrossile usando NADPH. In contrasto con l’intermedio idrossilato dell’ossidazione beta, l’intermedio beta qui è in configurazione D. Successivamente, l’acqua viene rimossa dai carboni 2 e 3 dell’intermedio idrossile per produrre una molecola trans a doppio legame. Infine, il doppio legame viene idrogenato per produrre un intermedio saturo. Il processo si ripete ciclicamente con l’aggiunta di un altro malonyl-ACP alla catena in crescita, finché alla fine viene prodotto un intermedio con 16 carboni (palmitoyl-CoA). A questo punto, la sintesi citoplasmatica cessa.
Enzimi della sintesi degli acidi grassi
L’acetil-CoA carbossilasi, che catalizza la sintesi del malonil-CoA, è l’unico enzima regolato nella sintesi degli acidi grassi. La sua regolazione coinvolge sia il controllo allosterico che la modifica covalente. L’enzima è noto per essere fosforilato sia da AMP Kinase che da Protein Kinase A. La defosforilazione è stimolata dalle fosfatasi attivate dal legame con l’insulina. La defosforilazione attiva l’enzima e favorisce il suo assemblaggio in un lungo polimero, mentre la fosforilazione inverte il processo. Il Palmitoyl-CoA lo inattiva allostericamente.
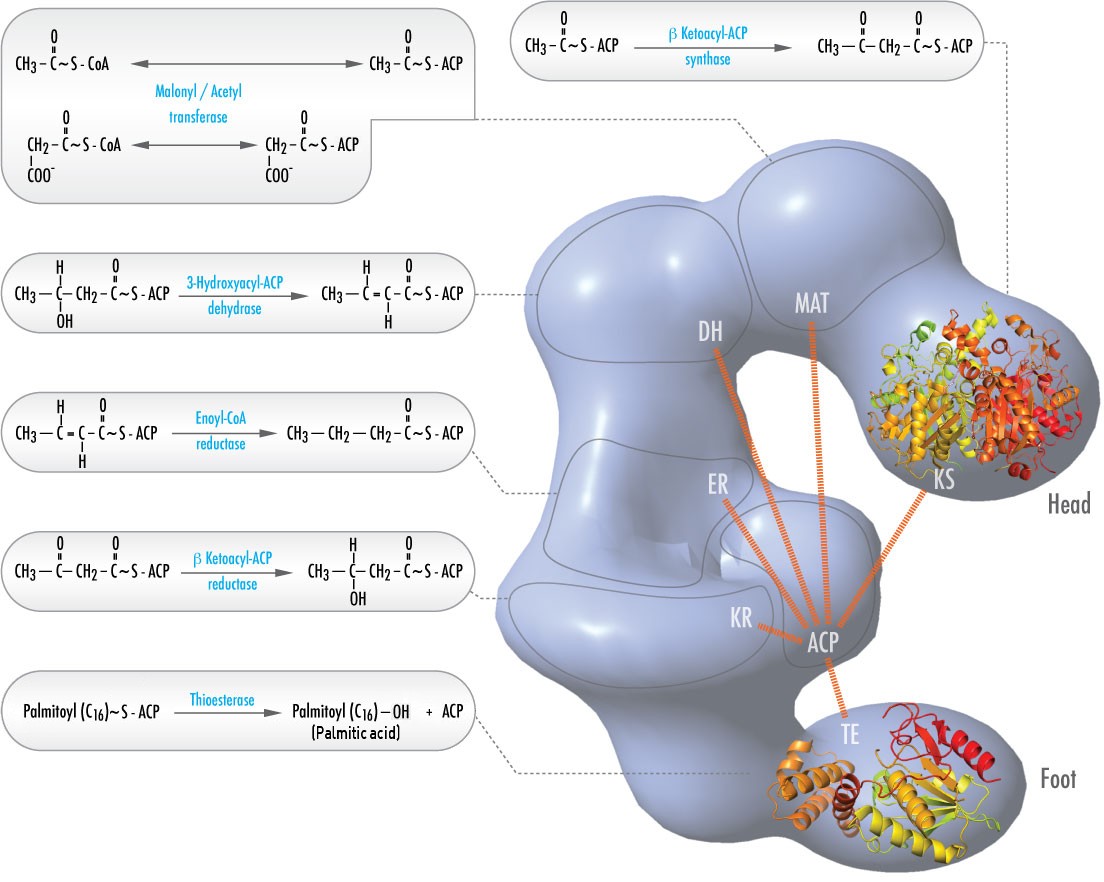
Negli animali, sei diverse attività catalitiche necessarie per le restanti azioni catalitiche per rendere completamente il palmitoil-CoA sono contenute in un unico complesso chiamato Sintasi degli Acidi Grassi (Figura \(\PageIndex{2}). Queste includono le transacilasi per scambiare il CoA con l’ACP sull’acetil-CoA e il malonil-CoA; una sintasi per catalizzare l’aggiunta dell’unità di due carboni dal malonil-ACP a tre carboni nel primo passo del processo di allungamento; una reduttasi per ridurre il chetone; una disidrasi per catalizzare la rimozione dell’acqua, e una reduttasi per ridurre il doppio legame trans. Nei batteri, queste attività si trovano su enzimi separati e non fanno parte di un complesso.
Allungamento degli acidi grassi
L’allungamento per rendere gli acidi grassi più lunghi di 16 carboni avviene nel reticolo endoplasmatico ed è catalizzato da enzimi definiti elongasi. Anche i mitocondri possono allungare gli acidi grassi, ma i loro materiali di partenza sono generalmente più corti di 16 carboni. I meccanismi in entrambi gli ambienti sono simili a quelli del citoplasma (un gruppo malonilico è usato per aggiungere due carboni, per esempio), ma il CoA è attaccato agli intermedi, non l’ACP. Inoltre, mentre la sintesi citoplasmatica impiega il complesso dell’acido grasso sintasi (Figura \(\PageIndex{2})), gli enzimi in questi organelli sono separabili e non fanno parte di un complesso.
Desaturazione degli acidi grassi
Gli acidi grassi sono sintetizzati in forma satura e la desaturazione avviene più tardi. Gli enzimi chiamati desaturasi catalizzano la formazione di doppi legami cis negli acidi grassi maturi. Questi enzimi si trovano nel reticolo endoplasmatico. Gli animali sono limitati negli acidi grassi desaturati che possono produrre, a causa dell’incapacità di catalizzare le reazioni oltre i carboni 9 e 10. Così, gli esseri umani possono produrre acido oleico, ma non possono sintetizzare acido linoleico o acido linolenico. Di conseguenza, questi due devono essere forniti nella dieta e sono indicati come acidi grassi essenziali.
Contribuenti
-
Dr. Kevin Ahern e Dr. Indira Rajagopal (Oregon State University)