Valutazione | Biopsicologia | Comparativa |Cognitiva | Sviluppo | Linguaggio | Differenze individuali | Personalità | Filosofia | Sociale |
Metodi | Statistica |Clinica | Educativa | Industriale |Professionale articoli | Psicologia mondiale |
Biologico:Genetica comportamentale – Psicologia evolutiva – Neuroanatomia – Neurochimica – Neuroendocrinologia – Neuroscienze – Psiconeuroimmunologia – Psicologia fisiologica – Psicofarmacologia (Indice, Outline)
| Rod cellula | |
|---|---|
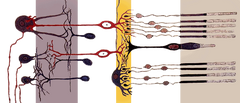
|
|
| Localizzazione | Retina |
| Funzione | Fotorecettore a bassa luminosità |
| Morfologia | a forma di asta |
| Connessioni presinaptiche | Nessuno |
| Connessioni postsinaptiche | Cellule bipolari e cellule orizzontali |
Le cellule Rod, o bastoncelli, sono cellule fotorecettrici nella retina dell’occhio che possono funzionare in una luce meno intensa rispetto all’altro tipo di fotorecettore, le cellule coniche. Poiché sono più sensibili alla luce, i bastoncelli sono responsabili della visione notturna. Chiamati così per la loro forma cilindrica, i bastoncelli sono concentrati ai bordi esterni della retina e sono usati nella visione periferica. Ci sono circa 120 milioni di cellule di bastoncelli nella retina umana.
Una cellula di bastoncello è abbastanza sensibile da rispondere a un singolo fotone di luce, ed è circa 100 volte più sensibile a un singolo fotone rispetto ai coni. Poiché i bastoncelli richiedono meno luce per funzionare rispetto ai coni, sono quindi la fonte primaria di informazioni visive durante la notte (visione scotopica). Le cellule del cono, d’altra parte, richiedono da decine a centinaia di fotoni per essere attivate. Inoltre, più cellule del bastoncello convergono su un singolo interneurone, raccogliendo e amplificando i segnali. Tuttavia, questa convergenza ha un costo per l’acuità visiva (o risoluzione dell’immagine) perché le informazioni messe insieme da più cellule sono meno distinte di quanto lo sarebbero se il sistema visivo ricevesse informazioni da ogni cellula del bastoncello individualmente. La convergenza delle cellule dei bastoncelli tende anche a rendere la visione periferica molto sensibile al movimento, ed è responsabile del fenomeno di un individuo che vede qualcosa di vago con la coda dell’occhio.
I bastoncelli sono un po’ più stretti dei coni ma hanno la stessa base strutturale. Il pigmento si trova sul lato esterno, adagiato sull’epitelio pigmentato. Questa estremità contiene molti dischi impilati, probabilmente dal ripiegamento verso l’interno della membrana limitante che circonda questa sezione, consentendo una superficie maggiore per il pigmento visivo e aumentando l’efficienza dell’assorbimento della luce. Poiché hanno solo un tipo di pigmento sensibile alla luce, piuttosto che i tre tipi che hanno le cellule coniche umane, i bastoncelli hanno poco, se non nessun, ruolo nella visione dei colori. Mentre questo rende i bastoncelli più sensibili a piccole quantità di luce, significa anche che la loro capacità di percepire i cambiamenti temporali, come le immagini che cambiano rapidamente, è meno accurata di quella dei coni.
Sperimenti di George Wald e altri hanno dimostrato che i bastoncelli sono più sensibili alle lunghezze d’onda della luce intorno ai 498 nm (verde-blu), e sono completamente insensibili alle lunghezze d’onda più lunghe di circa 640 nm (rosso). Questo fatto è responsabile dell’effetto Purkinje, in cui i colori blu appaiono più intensi rispetto ai rossi al crepuscolo, quando i bastoncelli subentrano come cellule responsabili della visione.
Come i coni, le cellule dei bastoncelli hanno un terminale sinaptico, un segmento interno e un segmento esterno. Il terminale sinaptico forma una sinapsi con un altro neurone, per esempio una cellula bipolare. Il segmento interno e quello esterno sono collegati da un cilium. Il segmento interno contiene organelli e il nucleo della cellula, mentre il segmento esterno della verga (abbreviato in ROS), che è rivolto verso la parte posteriore dell’occhio, contiene i materiali che assorbono la luce.
Risposta alla luce
Anatomia di una cellula a bastoncello
L’attivazione di una singola molecola di rodopsina, il pigmento fotosensibile dei bastoncelli, può portare a una grande reazione nella cellula perché il segnale viene amplificato. Una volta attivata, la rodopsina può attivare centinaia di molecole di trasducina, ognuna delle quali a sua volta attiva una molecola di fosfodiesterasi, che può scomporre oltre mille molecole di cGMP al secondo. Così i bastoncelli possono avere una grande risposta ad una piccola quantità di luce.
Poiché la componente retinica della rodopsina deriva dalla vitamina A, una carenza di vitamina A causa un deficit nel pigmento necessario alle cellule dei bastoncelli. Di conseguenza, un minor numero di cellule di bastoncello è in grado di rispondere sufficientemente in condizioni di oscurità, e poiché le cellule del cono sono scarsamente adattate per la vista al buio, si può arrivare alla cecità. Questa è la cecità notturna.
Ritorno allo stato di riposo
I bastoncelli fanno uso di tre meccanismi inibitori (meccanismi di feedback negativo) per permettere un rapido ritorno allo stato di riposo dopo un flash di luce.
In primo luogo, esiste una rodopsina chinasi (RK) che fosforilerebbe la coda citosolica della rodopsina attivata sulle serine multiple, inibendo parzialmente l’attivazione della transducina. Inoltre, una proteina inibitoria – la arrestina – si lega poi alle rodopsine fosforilate per inibire ulteriormente l’attività della rodopsina.
Mentre la arrestina spegne la rodopsina, una proteina RGS (che funziona come una proteina attivatrice di GTPasi (GAPs)) spinge la trasducina (proteina G) in uno stato “off” aumentando il tasso di idrolisi del GTP legato al GDP.
Siccome i canali sensibili al cGMP permettono non solo l’afflusso di ioni sodio, ma anche di ioni calcio, con la diminuzione della concentrazione di cGMP, i canali sensibili al cGMP vengono chiusi e si riduce il normale afflusso di ioni calcio. La diminuzione della concentrazione di ioni calcio stimola le proteine sensibili agli ioni calcio, che attiverebbero poi la guanilciclasi per ricostituire il cGMP, ripristinando rapidamente la sua concentrazione originale. Il ripristino apre i canali sensibili al cGMP e causa una depolarizzazione della membrana plasmatica.
Desensibilizzazione
Quando i bastoncelli sono esposti a un’alta concentrazione di fotoni per un periodo prolungato, diventano desensibilizzati (adattati) all’ambiente.
Quando la rodopsina è fosforilata dalla rodopsina chinasi (un membro delle GPCR chinasi (GRKs)), si lega con alta affinità all’arrestina. L’arrestina legata può contribuire al processo di desensibilizzazione in almeno due modi. In primo luogo, impedisce l’interazione tra la proteina G e il recettore attivato. In secondo luogo, serve come proteina adattatrice per aiutare il recettore al macchinario di endocitosi clatrina-dipendente (per indurre l’endocitosi mediata dal recettore).
Tabella
Confronto tra cellule di bastoncello e cono, da Kandel et al. (2000).
| Rods | Cones | |
|---|---|---|
| utilizzato per la visione notturna | utilizzato per la visione diurna | |
| molto sensibile alla luce; sensibile alla luce diffusa | non molto sensibile alla luce; sensibile solo alla luce diretta | |
| la perdita causa cecità notturna | la perdita causa cecità legale | |
| acuità visiva bassa | acuità visiva alta; migliore risoluzione spaziale | |
| non presente nella fovea | concentrata nella fovea | |
| risposta lenta, alla luce, stimoli aggiunti nel tempo | risposta veloce alla luce, possono percepire cambiamenti più rapidi negli stimoli | |
| hanno più pigmento dei coni, quindi possono rilevare meno luce | hanno meno pigmento dei bastoncelli, richiedono più luce per rilevare le immagini | |
| le pile di dischi di membranadischi chiusi non sono attaccati alla membrana cellulare | i dischi sono attaccati alla membrana esterna | |
| 20 volte più bastoncini che coni nella retina | ||
| un tipo di pigmento fotosensibile | tre tipi di pigmento fotosensibile negli esseri umani | |
| conferiscono la visione acromatica | conferiscono la visione a colori |
Riferimento
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principi di scienza neurale, 4a ed., pp.507-513. McGraw-Hill, New York.
- Fisiologia umana e meccanismi di malattia di Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Biologia molecolare della cellula, 5° ed., pp.919-921. Scienza Garland. Citare errore: Invalid
<ref>tag; name “Alberts” defined multiple times with different content
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principi di scienza neurale, 4a ed., pp.507-513. McGraw-Hill, New York.
.