La synthèse des acides gras se produit dans le cytoplasme et le réticulum endoplasmique de la cellule et est chimiquement similaire au processus de bêta-oxydation, mais avec quelques différences clés. La première d’entre elles a trait à la préparation des substrats pour les réactions qui produisent l’acide gras. Le transport de l’acétyl-CoA depuis la mitochondrie se produit lorsqu’il commence à s’accumuler. Deux molécules peuvent jouer un rôle dans son déplacement vers le cytoplasme : le citrate et l’acétylcarnitine. La jonction de l’oxaloacétate avec l’acétyl-CoA dans la mitochondrie crée du citrate qui traverse la membrane, suivi par l’action de la citrate lyase dans le cytoplasme de la cellule pour libérer l’acétyl-CoA et l’oxaloacétate. En outre, lorsque l’acétyl-CoA libre s’accumule dans la mitochondrie, il peut se combiner avec la carnitine et être transporté vers le cytoplasme.
En partant de deux acétyl-CoA, l’un est converti en malonyl-CoA par carboxylation catalysée par l’enzyme acétyl-CoA carboxylase (ACC), la seule enzyme régulatrice de la synthèse des acides gras (figure \(\PageIndex{1}\)). Ensuite, les deux molécules voient leurs portions CoA remplacées par une protéine porteuse appelée ACP (acyl-carrier protein) pour former l’acétyl-ACP et le malonyl-ACP. La jonction d’un acyl-ACP gras (dans ce cas, l’acétyl-ACP) avec le malonyl-ACP sépare le carboxyle qui a été ajouté et crée l’intermédiaire en haut à droite dans la figure de gauche.

Figure \(\PageIndex{1}\) : Synthèse des acides gras
À partir de ce moment, les réactions chimiques ressemblent à celles de la bêta-oxydation inversée. Tout d’abord, la cétone est réduite en un hydroxyle grâce au NADPH. Contrairement à l’intermédiaire hydroxylé de l’oxydation bêta, l’intermédiaire bêta est ici en configuration D. Ensuite, l’eau est retirée des carbones 2 et 3 de l’intermédiaire hydroxyle pour produire une molécule à double liaison trans. Enfin, la double liaison est hydrogénée pour donner un intermédiaire saturé. Le processus se répète avec l’ajout d’un autre malonyl-ACP à la chaîne croissante jusqu’à ce qu’un intermédiaire de 16 carbones soit finalement produit (palmitoyl-CoA). A ce stade, la synthèse cytoplasmique cesse.
Enzymes de la synthèse des acides gras
L’acétyl-CoA carboxylase, qui catalyse la synthèse du malonyl-CoA, est la seule enzyme régulée dans la synthèse des acides gras. Sa régulation implique à la fois un contrôle allostérique et une modification covalente. On sait que l’enzyme est phosphorylée à la fois par l’AMP Kinase et la protéine Kinase A. La déphosphorylation est stimulée par les phosphatases activées par la fixation de l’insuline. La déphosphorylation active l’enzyme et favorise son assemblage en un long polymère, tandis que la phosphorylation inverse le processus.Le citrate agit comme un activateur allostérique et peut également favoriser la polymérisation. Le palmitoyl-CoA l’inactive de manière allostérique.
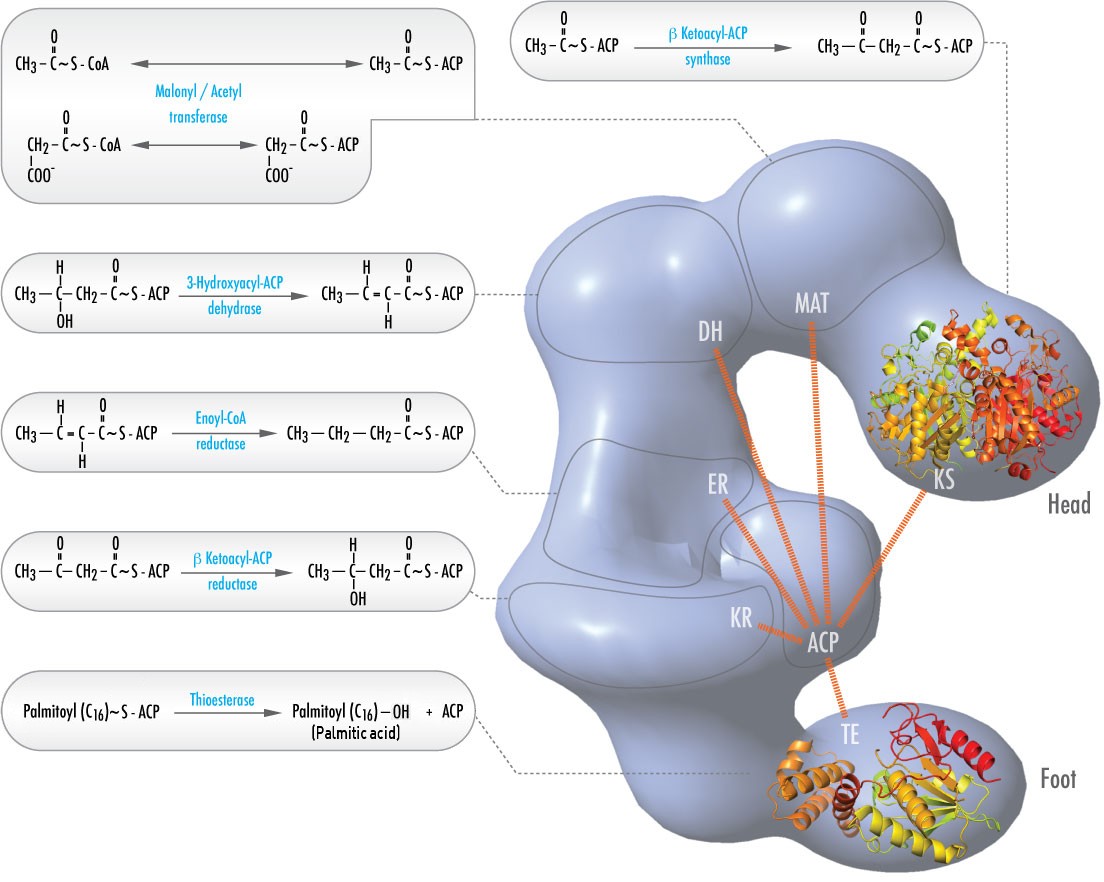
Chez les animaux, six activités catalytiques différentes nécessaires aux actions catalytiques restantes pour fabriquer entièrement le palmitoyl-CoA sont contenues dans un seul complexe appelé Fatty Acid Synthase (Figure \(\PageIndex{2}\)). Il s’agit notamment de transacylases pour échanger le CoA avec l’ACP sur l’acétyl-CoA et le malonyl-CoA ; d’une synthase pour catalyser l’addition de l’unité à deux carbones provenant du malonyl-ACP à trois carbones dans la première étape du processus d’élongation ; d’une réductase pour réduire la cétone ; d’une déshydrase pour catalyser l’élimination de l’eau et d’une réductase pour réduire la double liaison trans. Chez les bactéries, ces activités se trouvent sur des enzymes distinctes et ne font pas partie d’un complexe.
Élongation des acides gras
L’élongation pour rendre les acides gras plus longs que 16 carbones se produit dans le réticulum endoplasmique et est catalysée par des enzymes décrites comme des élongases. Les mitochondries peuvent également allonger les acides gras, mais leurs matériaux de départ sont généralement plus courts que 16 carbones. Les mécanismes dans ces deux environnements sont similaires à ceux du cytoplasme (un groupe malonyle est utilisé pour ajouter deux carbones, par exemple), mais le CoA est attaché aux intermédiaires, et non l’ACP. De plus, alors que la synthèse cytoplasmique emploie le complexe acide gras synthase (Figure \(\PageIndex{2}\)), les enzymes de ces organites sont séparables et ne font pas partie d’un complexe.
Désaturation des acides gras
Les acides gras sont synthétisés sous la forme saturée et la désaturation intervient plus tard. Des enzymes appelées désaturases catalysent la formation de doubles liaisons cis dans les acides gras matures. Ces enzymes se trouvent dans le réticulum endoplasmique. Les animaux sont limités dans les acides gras désaturés qu’ils peuvent fabriquer, en raison de leur incapacité à catalyser les réactions au-delà des carbones 9 et 10. Ainsi, les humains peuvent fabriquer de l’acide oléique, mais ne peuvent pas synthétiser l’acide linoléique ou l’acide linolénique. Par conséquent, ces deux derniers doivent être apportés par l’alimentation et sont appelés acides gras essentiels.
Contributeurs
-
Dr Kevin Ahern et Dr Indira Rajagopal (Oregon State University)
.