Objectifs d’apprentissage
- Nommer les composants qui génèrent les réponses sympathique et parasympathique du système nerveux autonome
- Expliquer les différences de sortie. connexions au sein des deux divisions du système nerveux autonome
- Décrire les molécules de signalisation et les protéines réceptrices impliquées dans la communication au sein des deux divisions du système nerveux autonome
Le système nerveux peut être divisé en deux parties fonctionnelles : le système nerveux somatique et le système nerveux autonome. Les principales différences entre ces deux systèmes sont évidentes dans les réponses que chacun produit. Le système nerveux somatique provoque la contraction des muscles squelettiques. Le système nerveux autonome contrôle les muscles cardiaques et lisses, ainsi que les tissus glandulaires. Le système nerveux somatique est associé aux réponses volontaires (bien que beaucoup puissent se produire sans conscience, comme la respiration), et le système nerveux autonome est associé aux réponses involontaires, comme celles liées à l’homéostasie.
Le système nerveux autonome régule de nombreux organes internes par un équilibre entre deux aspects, ou divisions. En ition au système endocrinien, le système nerveux autonome joue un rôle déterminant dans les mécanismes homéostatiques de l’organisme. Les deux divisions du système nerveux autonome sont la division sympathique et la division parasympathique. Le système sympathique est associé à la réaction de combat ou de fuite, et l’activité parasympathique est désignée par l’épithète de repos et de digestion. L’homéostasie est l’équilibre entre les deux systèmes. Au niveau de chaque effecteur cible, une double innervation détermine l’activité. Par exemple, le cœur reçoit des connexions des divisions sympathique et parasympathique. L’une entraîne une augmentation de la fréquence cardiaque, tandis que l’autre entraîne une diminution de la fréquence cardiaque.
Voyez cette vidéo pour en savoir plus sur l’adrénaline et la réaction de combat ou de fuite.
Lorsqu’on dit de quelqu’un qu’il a une poussée d’adrénaline, l’image des sauteurs à l’élastique ou des parachutistes vient généralement à l’esprit. Mais l’adrénaline, également connue sous le nom d’épinéphrine, est un produit chimique important dans la coordination de la réaction de combat ou de fuite du corps. Dans cette vidéo, vous pénétrez dans la physiologie de la réaction de lutte ou de fuite, telle qu’elle est envisagée pour un pompier. La réaction de son corps est le résultat de la division sympathique du système nerveux autonome qui provoque des changements à l’échelle du système alors qu’il se prépare à des réponses extrêmes. Quels sont les deux changements que l’adrénaline entraîne pour aider la réponse des muscles squelettiques ?
Division sympathique du système nerveux autonome
Pour répondre à une menace – se battre ou fuir – le système sympathique provoque des effets divergents, car de nombreux organes effecteurs différents sont activés ensemble dans un but commun. Une plus grande quantité d’oxygène doit être inhalée et délivrée aux muscles squelettiques. Les systèmes respiratoire, cardiovasculaire et musculo-squelettique sont tous activés ensemble. itionnellement, la transpiration empêche l’excès de chaleur provenant de la contraction musculaire de provoquer une surchauffe du corps. Le système digestif s’arrête pour que le sang n’absorbe pas de nutriments alors qu’il devrait fournir de l’oxygène aux muscles squelettiques. Pour coordonner toutes ces réponses, les connexions du système sympathique divergent d’une région limitée du système nerveux central (SNC) vers un large éventail de ganglions qui se projettent simultanément vers les nombreux organes effecteurs. L’ensemble complexe de structures qui composent la sortie du système sympathique permet à ces effecteurs disparates de se rassembler dans un changement coordonné et systémique.
La division sympathique du système nerveux autonome influence les différents systèmes organiques du corps par des connexions émergeant de la moelle épinière thoracique et lombaire supérieure. On parle de système thoracolombaire pour refléter cette base anatomique. Un neurone central dans la corne latérale de l’une de ces régions spinales se projette vers les ganglions adjacents à la colonne vertébrale par l’intermédiaire des racines spinales ventrales.
La majorité des ganglions du système sympathique appartiennent à un réseau de ganglions de la chaîne sympathique qui longe la colonne vertébrale. Les ganglions apparaissent comme une série d’amas de neurones reliés par des ponts axonaux. La chaîne compte généralement 23 ganglions de part et d’autre de la colonne vertébrale. Trois correspondent à la région cervicale, 12 à la région thoracique, quatre à la région lombaire et quatre à la région sacrée. Les niveaux cervical et sacré ne sont pas reliés à la moelle épinière directement par les racines spinales, mais par des connexions ascendantes ou descendantes à travers les ponts de la chaîne.
Un schéma qui montre les connexions du système sympathique est un peu comme un schéma de circuit qui montre les connexions électriques entre différents réceptacles et appareils. Dans la figure 1, les « circuits » du système sympathique sont intentionnellement simplifiés.

Figure 1. Connexions de la division sympathique du système nerveux autonome. Les neurones de la corne latérale de la moelle épinière (neurones préganglionnaires) se projettent vers les ganglions de la chaîne situés de part et d’autre de la colonne vertébrale ou vers les ganglions collatéraux (prévertébraux) qui sont antérieurs à la colonne vertébrale dans la cavité abdominale. Les axones de ces neurones ganglionnaires (fibres postganglionnaires) se projettent ensuite vers des effecteurs cibles dans tout le corps.
Pour poursuivre l’analogie du schéma de circuit, il existe trois types différents de « jonctions » qui opèrent au sein du système sympathique (figure 2). Le premier type est le plus direct : le nerf sympathique se projette sur le ganglion de la chaîne au même niveau que l’effecteur cible (l’organe, le tissu ou la glande à innerver).
Un exemple de ce type est le nerf spinal T1 qui fait synapse avec le ganglion de la chaîne T1 pour innerver la trachée. Les fibres de cette branche sont appelées rami communicantes blanches (singulier = ramus communicans) ; elles sont myélinisées et donc désignées comme blanches (voir figure 2a). L’axone du neurone central (la fibre préganglionnaire représentée en trait plein) fait synapse avec le neurone ganglionnaire (avec la fibre postganglionnaire représentée en trait pointillé). Ce neurone se projette ensuite vers un effecteur cible – dans ce cas, la trachée – par l’intermédiaire des rami communicantes grises, qui sont des axones non myélinisés.
Dans certains cas, les effecteurs cibles sont situés au-dessus ou en dessous du segment spinal où émerge la fibre préganglionnaire. En ce qui concerne le « câblage » impliqué, la synapse avec le neurone ganglionnaire se produit au niveau des ganglions de la chaîne supérieurs ou inférieurs à l’emplacement du neurone central. Le nerf spinal T1 qui innerve l’œil en est un exemple. Le nerf spinal remonte la chaîne jusqu’à ce qu’il atteigne le ganglion cervical supérieur, où il fait synapse avec le neurone postganglionnaire (voir figure 2b). Les ganglions cervicaux sont appelés ganglions paravertébraux, étant donné leur emplacement adjacent aux ganglions prévertébraux dans la chaîne sympathique.
Tous les axones des neurones centraux ne se terminent pas dans les ganglions de la chaîne. es branches partielles de la racine nerveuse ventrale continuent à travers la chaîne et sur l’un des ganglions collatéraux comme le nerf splanchnique supérieur ou le nerf splanchnique inférieur. Par exemple, le nerf splanchnique supérieur au niveau de T5 fait synapse avec un ganglion collatéral à l’extérieur de la chaîne avant d’établir la connexion avec les nerfs postganglionnaires qui innervent l’estomac (voir Figure 2c).
Les ganglions collatéraux, également appelés ganglions prévertébraux, sont situés en avant de la colonne vertébrale et reçoivent des entrées des nerfs splanchniques ainsi que des neurones sympathiques centraux. Ils sont associés au contrôle des organes de la cavité abdominale et sont également considérés comme faisant partie du système nerveux entérique. Les trois ganglions collatéraux sont le ganglion cœliaque, le ganglion mésentérique supérieur et le ganglion mésentérique inférieur (voir figure 1). Le mot cœliaque est dérivé du mot latin » coelom « , qui désigne une cavité corporelle (dans ce cas, la cavité abdominale), et le mot mésentérique désigne le système digestif.

Figure 2. Connexions sympathiques et ganglions en chaîne. L’axone d’un neurone sympathique central de la moelle épinière peut se projeter vers la périphérie de plusieurs façons différentes. (a) La fibre peut se projeter vers le ganglion au même niveau et faire une synapse sur un neurone ganglionnaire. (b) Une branche peut se projeter vers un ganglion plus supérieur ou inférieur dans la chaîne. (c) Une branche peut se projeter à travers le ramus communicans blanc, mais ne pas se terminer sur un neurone ganglionnaire de la chaîne. Au lieu de cela, elle se projette à travers l’un des nerfs splanchniques vers un ganglion collatéral ou la médullaire surrénale (non illustré).
Un axone du neurone central qui se projette vers un ganglion sympathique est appelé fibre ou neurone préganglionnaire, et représente la sortie du SNC vers le ganglion. Comme les ganglions sympathiques sont adjacents à la colonne vertébrale, les fibres sympathiques préganglionnaires sont relativement courtes et myélinisées. Une fibre postganglionnaire – l’axone d’un neurone ganglionnaire qui se projette vers l’effecteur cible – représente la sortie d’un ganglion qui influence directement l’organe.
Par rapport aux fibres préganglionnaires, les fibres sympathiques postganglionnaires sont longues en raison de la distance relativement plus grande entre le ganglion et l’effecteur cible. Ces fibres ne sont pas myélinisées. (Notez que le terme « neurone postganglionnaire » peut être utilisé pour décrire la projection d’un ganglion vers la cible. Le problème avec cet usage est que le corps cellulaire se trouve dans le ganglion, et que seule la fibre est postganglionnaire. Généralement, le terme neurone s’applique à la cellule entière.)
Un type de fibre sympathique préganglionnaire ne se termine pas dans un ganglion. Il s’agit des axones des neurones sympathiques centraux qui se projettent vers la médullaire surrénale, la partie intérieure de la glande surrénale. Ces axones sont toujours appelés fibres préganglionnaires, mais leur cible n’est pas un ganglion. La médullosurrénale libère des molécules de signalisation dans la circulation sanguine, plutôt que d’utiliser des axones pour communiquer avec les structures cibles. Les cellules de la médullosurrénale qui sont contactées par les fibres préganglionnaires sont appelées cellules chromaffines. Ces cellules sont des cellules neurosécrétoires qui se développent à partir de la crête neurale en même temps que les ganglions sympathiques, ce qui renforce l’idée que la glande est, fonctionnellement, un ganglion sympathique.
Les projections de la division sympathique du système nerveux autonome divergent largement, ce qui entraîne une large influence du système dans tout le corps. En réponse à une menace, le système sympathique augmenterait le rythme cardiaque et la fréquence respiratoire et provoquerait une augmentation du flux sanguin vers les muscles squelettiques et une diminution du flux sanguin vers le système digestif. La sécrétion des glandes sudoripares devrait également augmenter dans le cadre d’une réponse intégrée.
Tous ces changements physiologiques vont devoir se produire ensemble pour fuir la lionne en chasse, ou son équivalent moderne. Cette divergence se voit dans les schémas de ramification des neurones sympathiques préganglionnaires – un seul neurone sympathique préganglionnaire peut avoir 10 à 20 cibles. Un axone qui quitte un neurone central de la corne latérale dans la moelle épinière thoracolombaire va passer par le ramus communicans blanc et entrer dans la chaîne sympathique, où il se ramifiera vers une variété de cibles. Au niveau de la moelle épinière où la fibre sympathique préganglionnaire sort de la moelle épinière, une branche fera synapse sur un neurone du ganglion de chaîne adjacent.
Certaines branches s’étendront vers le haut ou vers le bas à un niveau différent des ganglions de chaîne. D’autres branches traverseront les ganglions de la chaîne et se projetteront à travers l’un des nerfs splanchniques vers un ganglion collatéral. Enfin, certaines branches peuvent se projeter à travers les nerfs splanchniques vers la médullaire surrénale. Toutes ces ramifications signifient qu’un neurone préganglionnaire peut influencer très largement différentes régions du système sympathique, en agissant sur des organes largement distribués.
Division parasympathique du système nerveux autonome
La division parasympathique du système nerveux autonome est nommée ainsi parce que ses neurones centraux sont situés de part et d’autre de la région thoracolombaire de la moelle épinière (para- = « à côté » ou « près »). Le système parasympathique peut également être appelé système craniosacré (ou exutoire) parce que les neurones préganglionnaires sont situés dans les noyaux du tronc cérébral et de la corne latérale de la moelle épinière sacrée.
Les connexions, ou « circuits », de la division parasympathique sont similaires à la disposition générale de la division sympathique avec quelques différences spécifiques (figure 3). Les fibres préganglionnaires de la région crânienne circulent dans les nerfs crâniens, tandis que les fibres préganglionnaires de la région sacrée circulent dans les nerfs spinaux. Les cibles de ces fibres sont les ganglions terminaux, qui sont situés à proximité, voire à l’intérieur, de l’effecteur cible. Ces ganglions sont souvent appelés ganglions intramuraux lorsqu’ils se trouvent dans les parois de l’organe cible. La fibre postganglionnaire se projette depuis les ganglions terminaux sur une courte distance vers l’effecteur cible ou vers le tissu cible spécifique à l’intérieur de l’organe. Si l’on compare les longueurs relatives des axones dans le système parasympathique, les fibres préganglionnaires sont longues et les fibres postganglionnaires sont courtes car les ganglions sont proches des effecteurs cibles – et parfois à l’intérieur de ceux-ci.
La composante crânienne du système parasympathique est basée dans des noyaux particuliers du tronc cérébral. Dans le mésencéphale, le noyau d’Eddinger-Westphal fait partie du complexe oculomoteur, et les axones de ces neurones voyagent avec les fibres du nerf oculomoteur (nerf crânien III) qui innervent les muscles extra-oculaires. Les fibres parasympathiques préganglionnaires du nerf crânien III se terminent dans le ganglion ciliaire, qui est situé dans l’orbite postérieure. Les fibres parasympathiques postganglionnaires se projettent ensuite sur le muscle lisse de l’iris pour contrôler la taille de la pupille. Dans la partie supérieure de la moelle, les noyaux salivaires contiennent des neurones dont les axones se projettent par les nerfs faciaux et glossopharyngiens vers les ganglions qui contrôlent les glandes salivaires. La production de larmes est influencée par les fibres parasympathiques du nerf facial, qui activent un ganglion, et finalement la glande lacrymale (larme).
Les neurones du noyau dorsal du nerf vague et du noyau ambigu se projettent par le nerf vague (nerf crânien X) vers les ganglions terminaux des cavités thoracique et abdominale. Les fibres préganglionnaires parasympathiques influencent principalement le cœur, les bronches et l’œsophage dans la cavité thoracique et l’estomac, le foie, le pancréas, la vésicule biliaire et l’intestin grêle dans la cavité abdominale. Les fibres postganglionnaires des ganglions activés par le nerf vague sont souvent incorporées à la structure de l’organe, comme le plexus mésentérique des organes du tube digestif et les ganglions intramuraux.

Figure 3. Connexions de la division parasympathique du système nerveux autonome Les neurones provenant des noyaux du tronc cérébral, ou de la corne latérale de la moelle épinière sacrée, se projettent vers des ganglions terminaux à proximité ou à l’intérieur des différents organes du corps. Les axones de ces neurones ganglionnaires projettent ensuite la courte distance vers ces effecteurs cibles.
Signalisation chimique dans le système nerveux autonome
Lorsqu’un neurone autonome se connecte à une cible, il y a une synapse. Le signal électrique du potentiel d’action provoque la libération d’une molécule de signalisation, qui va se lier à des protéines réceptrices sur la cellule cible. Les synapses du système autonome sont classées comme étant soit cholinergiques, ce qui signifie que l’acétylcholine (ACh) est libérée, soit adrénergiques, ce qui signifie que la norépinéphrine est libérée. Les termes cholinergique et adrénergique font référence non seulement à la molécule de signalisation qui est libérée, mais aussi à la classe de récepteurs à laquelle chacune se lie.
Le système cholinergique comprend deux classes de récepteurs : le récepteur nicotinique et le récepteur muscarinique. Les deux types de récepteurs se lient à l’ACh et provoquent des changements dans la cellule cible. Le récepteur nicotinique est un canal cationique à libération par ligand et le récepteur muscarinique est un récepteur couplé à une protéine G. Les récepteurs sont nommés d’après leur nom. Les récepteurs sont nommés et différenciés par les autres molécules qui s’y lient. Alors que la nicotine se lie au récepteur nicotinique et que la muscarine se lie au récepteur muscarinique, il n’y a pas de réactivité croisée entre les récepteurs. La situation est similaire à celle des serrures et des clés.
Imaginez deux serrures – l’une pour une salle de classe et l’autre pour un bureau – qui sont ouvertes par deux clés distinctes. La clé de la salle de classe n’ouvrira pas la porte du bureau et la clé du bureau n’ouvrira pas la porte de la salle de classe. Ceci est similaire à la spécificité de la nicotine et de la muscarine pour leurs récepteurs. Cependant, une clé passe-partout peut ouvrir plusieurs serrures, par exemple une clé passe-partout pour le département de biologie qui ouvre à la fois la porte de la salle de classe et celle du bureau. Ceci est similaire à l’ACh qui se lie aux deux types de récepteurs. Les molécules qui définissent ces récepteurs ne sont pas cruciales – elles sont simplement des outils que les chercheurs peuvent utiliser en laboratoire. Ces molécules sont exogènes, ce qui signifie qu’elles sont fabriquées en dehors du corps humain, de sorte qu’un chercheur peut les utiliser sans résultats endogènes confondants (résultats causés par les molécules produites dans le corps).
Le système adrénergique possède également deux types de récepteurs, nommés récepteur alpha (α)-adrénergique et récepteur bêta (β)-adrénergique. Contrairement aux récepteurs cholinergiques, ces types de récepteurs ne sont pas classés en fonction des médicaments qui peuvent s’y lier. Ils sont tous des récepteurs couplés aux protéines G. Il existe trois types de récepteurs α-adrénergiques, appelés α1, α2 et α3, et deux types de récepteurs β-adrénergiques, appelés β1 et β2. Un aspect itionnel du système adrénergique est qu’il existe une seconde molécule de signalisation appelée épinéphrine. La différence chimique entre la norépinéphrine et l’épinéphrine est l’ition d’un groupe méthyle (CH3) dans l’épinéphrine. Le préfixe « nor- » fait en fait référence à cette différence chimique, dans laquelle il manque un groupe méthyle.
Le terme adrénergique devrait vous rappeler le mot adrénaline, associé à la réaction de combat ou de fuite décrite au début du chapitre. L’adrénaline et l’épinéphrine sont deux noms pour la même molécule. La glande surrénale (en latin, ad- = « sur le dessus » ; renal = « rein ») sécrète l’adrénaline. La terminaison « -ine » indique que la substance chimique est dérivée, ou extraite, de la glande surrénale. Une construction similaire à partir du grec au lieu du latin donne le mot épinéphrine (epi- = « au-dessus » ; nephr- = « rein »). Dans l’usage scientifique, l’épinéphrine est préférée aux États-Unis, tandis que l’adrénaline est préférée en Grande-Bretagne, car « adrénaline » était autrefois un nom de médicament enregistré et breveté aux États-Unis. Bien que ce médicament ne soit plus commercialisé, la convention consistant à désigner cette molécule par ces deux noms différents persiste. De même, la norépinéphrine et la noradrénaline sont deux noms pour la même molécule.
Ayant compris les systèmes cholinergique et adrénergique, leur rôle dans le système autonome est relativement simple à comprendre. Toutes les fibres préganglionnaires, tant sympathiques que parasympathiques, libèrent de l’ACh. Tous les neurones ganglionnaires – les cibles de ces fibres préganglionnaires – possèdent des récepteurs nicotiniques dans leurs membranes cellulaires. Le récepteur nicotinique est un canal cationique fixé par un ligand qui entraîne la dépolarisation de la membrane postsynaptique. Les fibres parasympathiques postganglionnaires libèrent également de l’ACh, mais les récepteurs de leurs cibles sont des récepteurs muscariniques, qui sont des récepteurs couplés aux protéines G et ne provoquent pas exclusivement la dépolarisation de la membrane postsynaptique. Les fibres sympathiques postganglionnaires libèrent de la noradrénaline, à l’exception des fibres qui se projettent sur les glandes sudoripares et sur les vaisseaux sanguins associés aux muscles squelettiques, qui libèrent de l’ACh (tableau 1).
| Tableau 1 | ||
|---|---|---|
| Sympathique | Parasympathique | |
| Preganglionnaire | Acétylcholine >récepteur nicotinique | Acétylcholine >. récepteur nicotinique | Postganglionnaire | Norépinéphrine > a ou B-adrénergiques Acétylcholine > récepteur muscarinique (associé aux glandes sudoripares et aux vaisseaux sanguins associés aux muscles squelettiques uniquement |
Acétylcholine > récepteur muscarinique |
Les molécules de signalisation peuvent appartenir à deux grands groupes. Les neurotransmetteurs sont libérés au niveau des synapses, tandis que les hormones sont libérées dans la circulation sanguine. Ce sont des définitions simplistes, mais elles peuvent aider à clarifier ce point. L’acétylcholine peut être considérée comme un neurotransmetteur car elle est libérée par les axones au niveau des synapses. Le système adrénergique, cependant, présente un défi. Les fibres sympathiques postganglionnaires libèrent de la norépinéphrine, qui peut être considérée comme un neurotransmetteur. Mais la médullosurrénale libère l’épinéphrine et la noradrénaline dans la circulation, elles doivent donc être considérées comme des hormones.
Ce que l’on appelle ici des synapses peut ne pas correspondre à la définition la plus stricte de la synapse. Certaines sources désigneront la connexion entre une fibre postganglionnaire et un effecteur cible comme des jonctions neuroeffectives ; les neurotransmetteurs, tels que définis ci-dessus, seraient appelés des neuromodulateurs. La structure des connexions postganglionnaires n’est pas le bulbe terminal synaptique typique que l’on trouve à la jonction neuromusculaire, mais plutôt des chaînes de renflements sur la longueur d’une fibre postganglionnaire appelée varicosité (Figure 4).
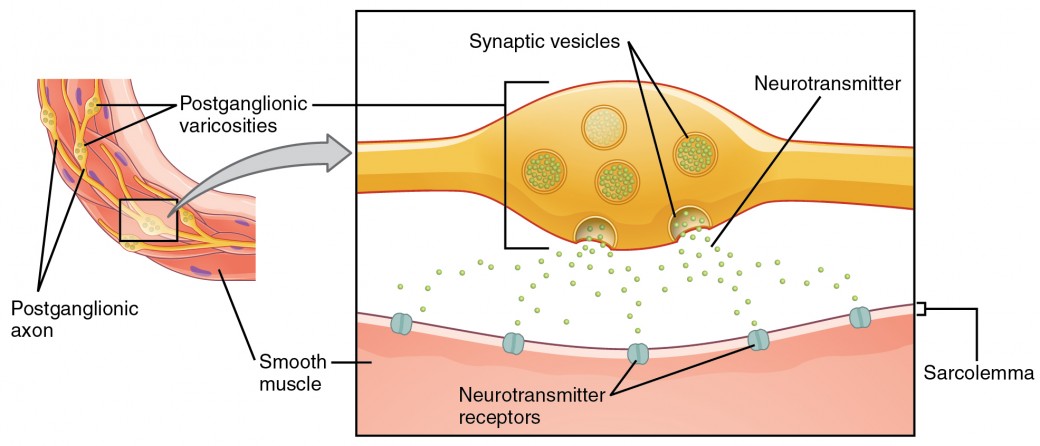
Figure 4. Varicosités autonomes. La connexion entre les fibres autonomes et les effecteurs cibles n’est pas la même que la synapse typique, comme la jonction neuromusculaire. Au lieu d’un bulbe terminal synaptique, un neurotransmetteur est libéré par des renflements sur la longueur d’une fibre qui établit un réseau étendu de connexions dans l’effecteur cible.
Connexions quotidiennes : Combattre ou fuir ? Et la peur et le gel ?
L’utilisation originale de l’épithète » combat ou fuite » provient d’un scientifique nommé Walter Cannon qui travaillait à Harvard en 1915. Le concept d’homéostasie et le fonctionnement du système sympathique avaient été introduits en France au siècle précédent. Cannon a développé cette idée et a introduit l’idée qu’un animal réagit à une menace en se préparant à se battre ou à s’enfuir. La nature de cette réponse a été expliquée en détail dans un livre sur la physiologie de la douleur, de la faim, de la peur et de la rage.
Lorsque les étudiants apprennent à connaître le système sympathique et la réaction de combat ou de fuite, ils s’arrêtent souvent pour s’interroger sur d’autres réponses. Si vous étiez confronté à une lionne courant vers vous, comme illustré au début de ce chapitre, vous enfuiriez-vous ou resteriez-vous debout ? Certaines personnes répondraient qu’elles se figeraient et ne sauraient pas quoi faire. Le système autonome ne fait-il pas plus que combattre, fuir, se reposer ou digérer ? Qu’en est-il de la peur et de la paralysie face à une menace ?
L’épithète courante de « combat ou fuite » est élargie pour devenir « combat, fuite ou frayeur » ou même « combat, fuite, frayeur ou gel ». La contribution originale de Cannon était une phrase accrocheuse pour exprimer une partie de ce que le système nerveux fait en réponse à une menace, mais elle est incomplète. Le système sympathique est responsable des réponses physiologiques aux états émotionnels. On peut dire que le nom » sympathique » signifie cela (sym- = » ensemble » ; -pathos = » douleur « , » souffrance » ou » émotion « ).
Voir cette vidéo pour en savoir plus sur le système nerveux.
Comme décrit dans cette vidéo, le système nerveux a une façon de gérer les menaces et le stress qui est distincte du contrôle conscient du système nerveux somatique. Ce système vient d’une époque où les menaces étaient liées à la survie, mais à l’époque moderne, ces réponses font partie du stress et de l’anxiété. Cette vidéo décrit comment le système autonome ne constitue qu’une partie de la réponse aux menaces, ou facteurs de stress. Quel autre système organique est impliqué, et quelle partie du cerveau coordonne les deux systèmes pour l’ensemble de la réponse, y compris l’épinéphrine (adrénaline) et le cortisol?
.