Évaluation | Biopsychologie | Comparatif |Cognitif | Développement | Langage | Différences individuelles. |Personnalité |Philosophie |Sociale |
Méthodes |Statistiques |Clinique |Education |Industrie |Eléments professionnels |Psychologie mondiale |
Biologique :Génétique comportementale – Psychologie évolutive – Neuroanatomie – Neurochimie – Neuroendocrinologie -Neuroscience – Psychoneuroimmunologie – Psychologie physiologique – Psychopharmacologie(Index, Outline)
| Rod cell | |
|---|---|
|
|
| Localisation | Rétine |
| Fonction | Photorécepteur de faible luminosité |
| Morphologie | en forme de bâtonnet |
| Connexions présynaptiques | Aucune |
| Connexions postsynaptiques | Cellules bipolaires et cellules horizontales |
Les cellules des bâtonnets, ou bâtonnets, sont des cellules photoréceptrices de la rétine de l’œil qui peuvent fonctionner dans une lumière moins intense que l’autre type de photorécepteur, les cellules coniques. Parce qu’ils sont plus sensibles à la lumière, les bâtonnets sont responsables de la vision nocturne. Appelés ainsi en raison de leur forme cylindrique, les bâtonnets sont concentrés sur les bords extérieurs de la rétine et sont utilisés pour la vision périphérique. Il existe environ 120 millions de cellules bâtonnets dans la rétine humaine.
Une cellule bâtonnet est suffisamment sensible pour répondre à un seul photon de lumière, et est environ 100 fois plus sensible à un seul photon que les cônes. Comme les bâtonnets ont besoin de moins de lumière pour fonctionner que les cônes, ils sont donc la principale source d’information visuelle la nuit (vision scotopique). Les cellules coniques, en revanche, ont besoin de dizaines ou de centaines de photons pour être activées. De plus, plusieurs cellules à bâtonnets convergent vers un seul interneurone, recueillant et amplifiant les signaux. Toutefois, cette convergence a un coût pour l’acuité visuelle (ou la résolution de l’image), car les informations regroupées de plusieurs cellules sont moins distinctes que si le système visuel recevait les informations de chaque cellule individuellement. La convergence des cellules à bâtonnets tend également à rendre la vision périphérique très sensible au mouvement, et est responsable du phénomène d’un individu qui voit quelque chose de vague se produire du coin de l’œil.
Les bâtonnets sont un peu plus étroits que les cônes mais ont la même base structurelle. Le pigment se trouve sur la face externe, reposant sur l’épithélium pigmentaire. Cette extrémité contient de nombreux disques empilés, probablement à cause du repli vers l’intérieur de la membrane limitante entourant cette section, ce qui permet d’obtenir une surface plus élevée pour le pigment visuel et d’augmenter l’efficacité de l’absorption de la lumière. Parce qu’ils ne possèdent qu’un seul type de pigment sensible à la lumière, plutôt que les trois types que possèdent les cellules coniques humaines, les bâtonnets ont peu, voire aucun, rôle dans la vision des couleurs.
Les bâtonnets répondent également plus lentement à la lumière que les cônes, de sorte que les stimuli qu’ils reçoivent s’additionnent sur environ 100 millisecondes. Si cela rend les bâtonnets plus sensibles à de plus petites quantités de lumière, cela signifie également que leur capacité à percevoir les changements temporels, comme les images qui changent rapidement, est moins précise que celle des cônes.
Des expériences menées par George Wald et d’autres ont montré que les bâtonnets sont plus sensibles aux longueurs d’onde de la lumière autour de 498 nm (vert-bleu), et sont complètement insensibles aux longueurs d’onde supérieures à environ 640 nm (rouge). Ce fait est responsable de l’effet Purkinje, dans lequel les couleurs bleues apparaissent plus intenses par rapport aux rouges au crépuscule, lorsque les bâtonnets prennent le relais comme cellules responsables de la vision.
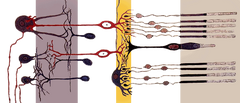
Comme les cônes, les cellules des bâtonnets possèdent un terminal synaptique, un segment interne et un segment externe. Le terminal synaptique forme une synapse avec un autre neurone, par exemple une cellule bipolaire. Les segments interne et externe sont reliés par un cilium. Le segment interne contient les organelles et le noyau de la cellule, tandis que le segment externe du bâtonnet (abrégé en ROS), qui pointe vers l’arrière de l’œil, contient les matériaux absorbant la lumière.
Réaction à la lumière
Anatomie d’une cellule de bâtonnet
L’activation d’une seule molécule de rhodopsine, le pigment photosensible des bâtonnets, peut entraîner une grande réaction dans la cellule car le signal est amplifié. Une fois activée, la rhodopsine peut activer des centaines de molécules de transducine, chacune d’entre elles activant à son tour une molécule de phosphodiestérase, qui peut décomposer plus de mille molécules de GMPc par seconde. Ainsi, les bâtonnets peuvent avoir une grande réponse à une petite quantité de lumière.
Comme le composant rétinien de la rhodopsine est dérivé de la vitamine A, une carence en vitamine A entraîne un déficit du pigment nécessaire aux cellules des bâtonnets. Par conséquent, moins de cellules à bâtonnets sont capables de répondre suffisamment dans des conditions plus sombres, et comme les cellules à cônes sont mal adaptées à la vue dans l’obscurité, la cécité peut en résulter. C’est l’héméralopie.
Retour à l’état de repos
Les bâtonnets font appel à trois mécanismes inhibiteurs (mécanismes de rétroaction négative) pour permettre un retour rapide à l’état de repos après un flash de lumière.
Premièrement, il existe une rhodopsine kinase (RK) qui phosphorerait la queue cytosolique de la rhodopsine activée sur les multiples sérines, inhibant partiellement l’activation de la transducine. De plus, une protéine inhibitrice – l’arrestine – se lie ensuite aux rhodopsines phosphorylées pour inhiber davantage l’activité de la rhodopsine.
Alors que l’arrestine éteint la rhodopsine, une protéine RGS (fonctionnant comme une protéine d’activation de la GTPase(GAPs)) entraîne la transducine (protéine G) dans un état « off » en augmentant le taux d’hydrolyse du GTP lié en GDP.
De même que les canaux sensibles au GMPc permettent non seulement l’influx d’ions sodium, mais aussi d’ions calcium, avec la diminution de la concentration de GMPc, les canaux sensibles au GMPc sont alors fermés et réduisent l’influx normal d’ions calcium. La diminution de la concentration d’ions calcium stimule les protéines sensibles aux ions calcium, qui activent alors la guanylyl cyclase pour reconstituer le GMPc, rétablissant rapidement sa concentration initiale. Cette restauration ouvre les canaux sensibles au GMPc et provoque une dépolarisation de la membrane plasmique.
Désensibilisation
Lorsque les bâtonnets sont exposés à une forte concentration de photons pendant une période prolongée, ils se désensibilisent (s’adaptent) à l’environnement.
Lorsque la rhodopsine est phosphorylée par la rhodopsine kinase (un membre des GPCR kinases(GRKs)), elle se lie avec une grande affinité à l’arrestine. L’arrestine liée peut contribuer au processus de désensibilisation d’au moins deux manières. Premièrement, elle empêche l’interaction entre la protéine G et le récepteur activé. Deuxièmement, elle sert de protéine adaptatrice pour aider le récepteur à la machinerie d’endocytose dépendante de la clathrine (pour induire une endocytose médiée par le récepteur).
Tableau
Comparaison des cellules de bâtonnets et de cônes, d’après Kandel et al. (2000).
| Rouges | Cônes |
|---|---|
| utilisé pour la vision nocturne | utilisé pour la vision diurne |
| très sensible à la lumière ; sensible à la lumière diffuse | pas très sensible à la lumière ; sensible uniquement à la lumière directe |
| La perte entraîne la cécité nocturne | La perte entraîne la cécité légale |
| Acuité visuelle faible | Acuité visuelle élevée ; meilleure résolution spatiale |
| concentrée dans la fovéa | |
| réponse lente, à la lumière, stimuli ajoutés au fil du temps | réponse rapide à la lumière, peuvent percevoir des changements plus rapides dans les stimuli |
| Possèdent plus de pigment que les cônes, donc peuvent détecter moins de lumière | Possèdent moins de pigment que les bâtonnets, nécessitent plus de lumière pour détecter les images |
| Les empilements de disques entourés de membrane-disques enfermés ne sont pas attachés à la membrane cellulaire | disques sont attachés à la membrane externe |
| 20 fois plus de bâtonnets que de cônes dans la rétine | |
| un type de pigment photosensible | trois types de pigment photosensible chez les humains. types de pigments photosensibles chez l’homme |
| confère une vision achromatique | confère une vision des couleurs |
Référence
- 1.0 1.1 1.2 1.3 Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principles of Neural Science, 4e édition, p.507-513. McGraw-Hill, New York.
- Physiologie humaine et mécanismes de la maladie par Arthur C. Guyton (1992) p.373
- 3.0 3.1 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter (2008). Biologie moléculaire de la cellule, 5e édition, p.919-921. Garland Science. Erreur de citation : Balise invalide
<ref>; nom « Alberts » défini plusieurs fois avec un contenu différent
- Kandel E.R., Schwartz, J.H., Jessell, T.M. (2000). Principes de la science des neurones, 4e édition, p.507-513. McGraw-Hill, New York.